
IX型コラーゲン破壊の診断
NC4ドメインのC末端部分に近い、IX型コラーゲンα1鎖のN末端部分の付近の新規な切断部位、及びNC3ドメインに近いCOL3ドメインにおける新規な結合部位で切断されたタンパク質断片の存在を検出又はモニターする方法。切断によって生じるネオエピトープ及びIX型コラーゲンに固有のNC4ドメインの切断N末端部分におけるエピトープに対するネオエピトープ抗体。このような方法の実施に有用な診断キット及び抗体も提示される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、血清及び滑液中の、IL−1が媒介するIX型コラーゲンの切断断片のレベルを決定することにより、関節疾患などの結合組織を冒す障害、とりわけ、変形性関節炎、関節リウマチ、反応性関節炎、腱断裂、骨粗しょう症、骨軟化症、骨折修復、及び動脈硬化を検出又はモニターすることを目的とした方法に関する。より詳細には、本発明は、IX型コラーゲンα1鎖のN末端部分の付近、又はより詳細にはNC4ドメインのC末端部分の近くにある新規な切断部位、及びNC3ドメインに近いCOL3ドメインにおける新規な切断部位で切断されたタンパク質断片の存在の検出又はモニタリングに関する。本発明はまた、切断により生じるネオエピトープ及びIX型コラーゲンに固有のNC4ドメインの切断N末端部分におけるエピトープに対するネオエピトープ抗体を開示するものである。このような方法の実施に有用な診断キット及び抗体も提示される。
【背景技術】
【0002】
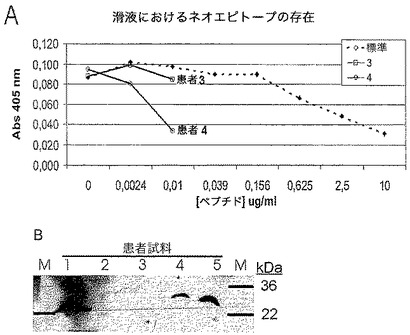
硝子軟骨のIX型コラーゲンには、主要な構成要素としてII型コラーゲンを有する線維網の、主要ではない構成要素が存在する。IX型コラーゲン分子は、ポリペプチド鎖α1、α2、及びα3からなるヘテロ三量体である(2)。それは、中断された三重らせんを有する原線維結合コラーゲン(FACIT)に属する。それぞれの鎖は、4個の非三重らせんドメインであるNC1、2、3、及び4に囲まれている3個の三重らせん(コラーゲン性)ドメインであるCOL1、2、及び3を含む(3)(図9も参照されたい)。ドメインの数字はC末端から数える。
【0003】
電子顕微鏡法を用いて、IX型コラーゲンがII型コラーゲン原線維の表面を装飾していること、及びNC4ドメインが球形構造を形成し、それが茎状のCOL3ドメインと共にII型コラーゲン原線維から突き出していることが示されている(4)。
【0004】
IX型コラーゲンは、II型コラーゲン分子及び他のIX型コラーゲン分子の両方への結合を介して、II型コラーゲン原線維に共有結合で架橋している(5〜7)。これらの結合は、ペプチド結合を切断しない作用物質による、成熟軟骨からのIX型コラーゲンの抽出を事実上不可能にする。
【0005】
NC4ドメインは、多くの分子、例えばヘパリン及び軟骨オリゴマーマトリックスタンパク質(COMP)に対する親和性を有することが示されているが(8〜10)、COLドメインはマトリリン−3と相互作用する(11)。相互作用するCOMP、MATN−3、及びCOL9における突然変異は、稀な遺伝性の、分布が不均一な疾患である多発性骨端異形成症を有する患者において見られており、骨端の成長及び骨化に影響する(12)。
【0006】
驚くべきことに、NC4断片は現在、軟骨破壊のバイオマーカーであることが示されている。切断は、軟骨の主要な構造的成分であるアグリカンが放出されてから、主要なコラーゲンが分解されるまでに生じる。X線観察のように破壊プロセスの後期段階の結果であるコラーゲンマーカーとは対照的に、NC4断片は、「引き返しの限界点」の前の破壊をもたらすプロセスのマーカーである。アグリカン関連マーカーは、分解進行の初期段階において、また組織の通常の適応化の際に放出され、したがって、病理学的プロセスを特別に示すものではない。さらに、アグリカンマーカーは炎症に関連することが多い。
【0007】
本発明に記載されているように、NC4の切断の後、II型コラーゲンに架橋したままであるIX型コラーゲンの大部分は、さらに切断されてその後の段階において放出される可能性を有している。本明細書における発明で開示されている、放出された断片の新たなC末端に対するネオエピトープ抗体は、このような切断を検出することができた。加えて、本発明において開示されているネオエピトープ抗血清を免疫組織化学で用いて、本明細書において報告されている新規な切断が軟骨のどこで(表面−深部、細胞周囲−領域間)生じているかを決定することができた。組織内に最初に残存していた断片の新たに形成されたN末端に対する抗体は、その後の段階におけるこのような断片の放出を検出することができた。タイミングについてのさらなる情報は、col3ドメインにおける切断により生じたネオエピトープを用いて得ることができた。多くの先行する刊行物及び特許出願が、IX型コラーゲンに特異的に結合するモノクローナル抗体を記載しているが、それらのいずれも、NC4付近の切断部位に特異的なネオエピトープ抗体については述べていない。
【0008】
特許出願WO1990008195A1は、ヒトIX型コラーゲン及びそれに由来する断片に免疫学的に結合するモノクローナル抗体の産生において抗原として有用な、ヒトIX型コラーゲンのセグメントに対応するオリゴペプチドを開示している。しかし、この特許出願において述べられているセグメントはIX型コラーゲンの病理学的破壊には関与しておらず、したがって、関節炎との関係においては期待されない。
【0009】
Inst of Nutraceutical Research PTYに譲渡されたWO2004110475A1において、発明は、関節炎の症状を有する個体に対する治療及び耐性誘導のための、NC4ドメインを含む組成物に関するものである。発明者らはまた、抗関節炎活性又は抗炎症活性を有するポリペプチドを結合組織から回収するための方法を記載している。上記の特許出願のように、抗体は、切断されていないIX型コラーゲンに対するものであり、ネオエピトープ抗体ではない。さらに、出願人らは、NC4を診断のためのマーカーとしては述べていない。
【0010】
Fibrogen Incに譲渡された特許出願WO1999021011A1は、血清中のIX型コラーゲンのレベルを決定することにより自己免疫障害及び結合組織障害を検出又はモニターするための方法を明らかにしている。IX型コラーゲンに対する抗体を調製する際に、NC4ドメインのN末端部分における配列が選択された。この抗体は、切断されていないIX型コラーゲンに対するものであり、本明細書の発明におけるような、固有の切断ネオエピトープに対するものではない。
【0011】
先行文献のいずれも、NC−4ドメインの排他的な放出をもたらす、このドメインにおいて生じる特定の切断部位については記載していない。組織が特定の疾患において完全に破壊されるという事実にも関わらず、切断の特徴は推測又は予測することができない。とりわけ配列の類似性の観点から、ウシ軟骨及びヒト軟骨において分子の切断が異なることも予想されなかった(図10)。
【0012】
したがって、特定の切断部位の発見は新規であり、新たな診断発明を開拓するものである。
【発明の概要】
【発明が解決しようとする課題】
【0013】
したがって、本発明者らは、滑液、血清、尿、及びあらゆる体液などのあらゆるヒト試料におけるIX型コラーゲン破壊の分析を容易にする、同定された新規な切断部位に対する方法として、高感度のウェスタンブロット技術及び高感度の阻害ELISAを開発した。この種のアッセイは、同定された切断が、患者における軟骨の分解及び関節の劣化において出現するかどうか、並びにいつ出現するかを明らかにするために用いることができた。本明細書の発明の特異的なネオエピトープ抗体を用いることで、アッセイを、疾患の進行の診断及び治療に対する疾患の応答のモニタリングに用いることができる。例えば、骨折の治療中のスポーツマンに対して、トレーニングプログラムを再開する最適な時期を決定するためである。
【課題を解決するための手段】
【0014】
インターロイキン−1誘導性のウシ鼻軟骨の分解を用いて、16日間にわたり組織における異化事象を研究した。培地を2D電気泳動(等電点電気泳動法及びSDS PAGE)によって分画化した。ペプチドマスフィンガープリンティングによる成分の同定により、12日目及び16日目に、IX型コラーゲンα1鎖のNC4ドメインなどのタンパク質断片の放出が明らかになった。
【0015】
NC4ドメインのN末端部分におけるエピトープに対する新規なペプチド抗体は所見を裏付けるものであり、既に9日目に、断片の1つが存在することを示した。
【0016】
2つの最も豊富な断片の質量分析により、最も小さい断片が、配列内のアルギニン−258とイソロイシン−259との間で切断されてC末端ETCNELPAR−(配列番号1)及びN末端−ITPGARSP(配列番号2)(ETCNELPAR258−COOH及びNH2−ITPGARSP)を生じさせる、ほぼ完全なNC4ドメインを含むことが明らかになった。
【0017】
大きい方の断片は、NC4ドメインと、NC3ドメインに近い、C末端RGPPGPPGPPGPSG−(配列番号3)及びN末端−TIGFHDGD(配列番号4)(RGPPGPPGPPGPSG400−COOH及び−NH2−TIGFHDGD)を生じさせる、COL3におけるグリシン−400とスレオニン−401との間に位置する切断部位を有するCOL3ドメインとを含む。複数の(IX型)コラーゲンα1のN末端配列の存在は、放出された分子が、IL−1処理の前に、又はその処理によって、元のN末端に非常に近い部位で切断されていることを示している。
【0018】
マトリックスメタロプロテイナーゼ13(MMP−13)がこの期間において活性であり、フィブロモジュリンを切断することが示されている(1)。MMP−13で処理した軟骨移植片は、IL−1処理で得られるものと同じサイズ及び同じ切断部位を有する(IX型)コラーゲンα1断片を放出することが示された。示されたデータは、MMP−13を伴うと考えられる潜在的に重要な分解事象、及び最終的なII型コラーゲンの損失に先行する分解事象を説明するものである。
【0019】
ヒト軟骨におけるIX型コラーゲンの切断のさらなる研究により、NC−4ドメインにヒトIX型コラーゲンα1鎖の特異的な切断部位もあったが、これは驚くべきことにウシコラーゲンのものとは異なる部位に生じたことが明らかになった。NC−4ドメインにおける、ヒトIX型コラーゲンα1鎖のCHELPARITPSQTTDERGPP(配列番号5)におけるグルタミン−263とスレオニン−264との間の部位における切断により、C末端CHELPARITPSQ−(配列番号6)及びN末端−TTDERGPP)(配列番号7)(−CHELPARITPSQ263−COOH及びNH2−TTDERGPP)が生じる。COL3ドメイン内の部位に相当する大きい方の断片は、ウシ及びヒトの軟骨に由来するもので同じサイズであり、ウシにおけるS399がヒトRGPPGPPGPPGPRG400TIGFHDGD408(配列番号8)においてはR399に交換されているにもかかわらず、切断は同じ部位、すなわちG400とT401との間で生じる。ヒトRGPPGPPGPPGPRG400TIGFHDGD408(配列番号8)の切断の結果、2つのネオエピトープRGPPGPPGPPGPRG(配列番号9)及びTIGFHDGD408(配列番号10)が生じる。ヒトNC−4及びCOL3の新たなネオエピトープ末端の全てに対する抗体が、標準的な方法を用いて開発され、切断断片との特異的な反応性を示した。抗体を用いて、エピトープを検出するためのELISAを開発した。これらのELISAを用いて、関節疾患などの結合組織を冒す障害、とりわけ、変形性関節炎、並びに関節リウマチ、腱断裂、骨折修復、及び動脈硬化を有する患者から得た滑液におけるエピトープの存在を示した。本発明の特徴は、本発明を限定するものとは解釈されない以下の実施例を参照することにより、より明瞭に理解されよう。
【図面の簡単な説明】
【0020】
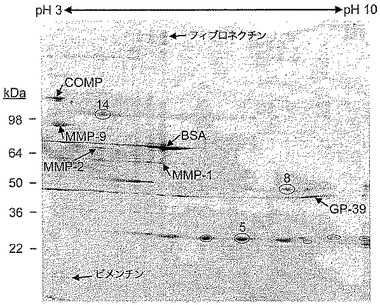
【図1】2D SDS PAGEを用いて分離した、16日目の、IL−1で刺激したウシ鼻軟骨培地から得たタンパク質を示す図である。一次元目である等電点電気泳動は3〜10のpH範囲にわたるものであり、二次元目は4〜20%の勾配ゲルを用いた。画像の左側の数字は参照マーカータンパク質の位置を示す。輪で印を付けたスポットは、(IX型)コラーゲンα1であると同定されたスポットを示す。異なるIX型コラーゲン断片は、本文中において、28kDa断片、50kDa断片、及び100kDa断片であるとみなされている。画像において、分析した特定のスポットの例に5、8、及び14と印を付けている。これらのスポットのMALDI−TOF MS(マトリックス支援レーザー脱離イオン化飛行時間型質量分析)の結果を、以下の図11a、b、及びcに示す。
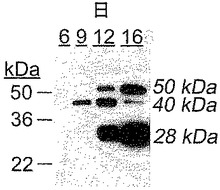
【図2】IL−1で刺激した軟骨移植片の、NC4抗血清を用いたウェスタンブロットを示す図である。IL−1で刺激した軟骨培養物の6日目、9日目、12日目、及び16日目の培地を、1D SDS PAGEゲル上で分離した。28kDa、40kDa、及び50kDaの見かけの質量での陽性シグナルが明瞭に視認可能である。
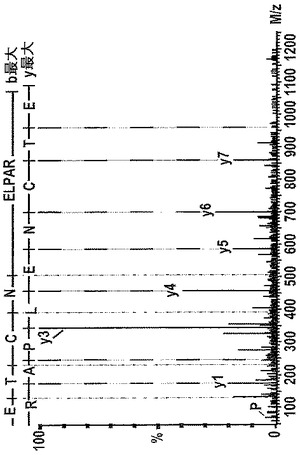
【図3】28kDaのNC4断片の最もC末端のペプチドから得たタンデム質量分析のデータを示す図である。28kDaの断片をAspNでゲル内消化し、ESI−Qtof MS(エレクトロスプレーイオン化−四重極飛行時間型質量分析)を用いてオンラインで逆相LC(液体クロマトグラフィー、C18カラム)で分析した。推定されるペプチド配列はETCNELPAR(配列番号1)である。IL−1で刺激した軟骨から得た培地(16日目)をまず、2D SDS PAGEで分離した。この注釈付きのスペクトルは、BioLynxソフトウェア(ペプチド配列決定、Waters Djupdalsvagen 12−14 Sollentuna Sweden)において生じたものであった。ペプチドのデルタ質量は<0.01Daであり、yイオンの平均誤差は0.01Daであり、強度の閾値は0.75%に設定した。
【図4a】NC4抗血清を用いた、IL−1及びMMP−13で処理した軟骨の培地のウェスタンブロットを示す図である。IL−1(16日目)又はMMP−13で処理したウシ鼻軟骨から放出されたタンパク質を16%SDS PAGEによって分離した。クマシーブルーG−250で染色したゲル上の同等の位置を切り取り、ゲル内消化し、ESI−IonTrap MS(エレクトロスプレーイオン化イオントラップ質量分析)を用いてオンラインで逆相クロマトグラフィー(C18カラム)で分析した。両方の28kDaの断片が、同一の最もC末端のペプチドETCNELPAR(配列番号1)を含んでいた(図3参照)。50kDaの断片は、P*がヒドロキシプロリンである、共通の最もC末端のペプチドRGPP*GPP*GPP*GPSG(配列番号3)を共有していた(図4b参照)。右側の挿入図は、40kDaのバンドを示す、同一のブロットの過剰曝露のバージョンである。
【図4b】ウシIX型コラーゲンの50kDaの断片における最もC末端のペプチドのタンデム質量分析を示す図である。50kDaの断片をトリプシンでゲル内消化し、IonTrap MSMSを用いてオンラインで逆相クロマトグラフィーで分析すると、最もC末端のペプチド(P*がヒドロキシプロリン残基であるRGPP*GPP*GPP*GPSG(配列番号3))のタンデム質量分析のデータが見られた。MMP−13消化溶液を16%SDS PAGEで分離し、クマシーブルーG−250で染色した。この注釈付きのスペクトルは、BioTools(Bruker Daltonics Scandinavia AB、Polygonvagen 79 Taby Sweden)において生じたものであった。許容誤差は0.2Daであり、bイオン及びyイオンの平均誤差は0.06Daであり、強度の閾値は10に設定した。
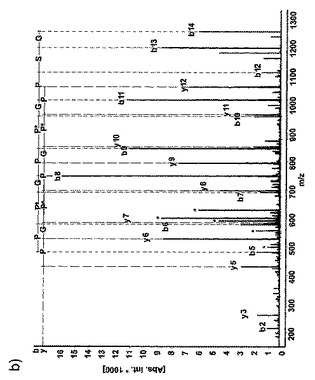
【図5a】MMP−13又はIL−1で処理したヒト軟骨の、NC4抗血清を用いたウェスタンブロットを示す図である。ヒト関節及びウシ鼻軟骨をMMP−13で消化し、後者はIL−1での刺激も行った(16日目)。消化溶液及び培地を16%ポリアクリルアミド−SDSゲル上で分離した。
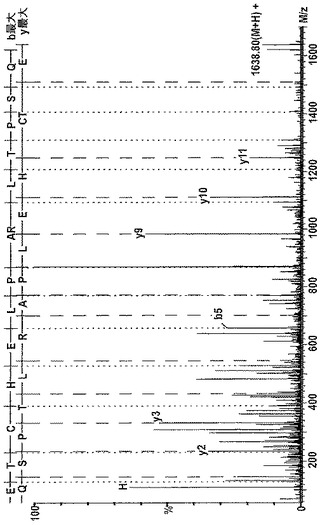
【図5b】ヒトNC4断片の最もC末端のペプチドのタンデム質量分析のデータを示す図である。断片をAsp−Nでゲル内消化し、LC−Qtof MSで分析した。ロイシン及びイソロイシンは正確な同一の質量を有しており、したがって、スペクトルにおけるLはL又はIに相当する。推定されるペプチド配列はETCHELPARITPSQ(配列番号11)である。MMP−13で消化した軟骨の培地を16%ポリアクリルアミド−SDSゲル上で分画化し、その後、質量分析を行った。
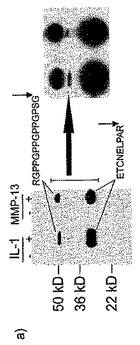
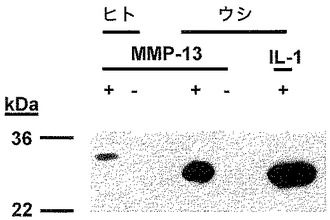
【図6】MMP−13で消化したヒト関節軟骨の培地の、ネオエピトープ抗血清を用いたウェスタンブロットを示す図である。ヒト関節軟骨をMMP−13で消化した。消化溶液を16%ポリアクリルアミド−SDSゲル上で分離し、電気泳動でニトロセルロース膜に移し、それを、全ての変異体を認識するNC4における内部エピトープに対するウサギ抗血清(右パネル)、及び左パネルの、PARITPSQ263に対するネオエピトープ抗体とインキュベートした。結合した抗体を、ペルオキシダーゼ結合抗ウサギIgG抗体で検出した。切断部位全体にわたる連続的な配列を含む大きい方の断片とのネオエピトープの反応性がないことは、切断断片に対する特異性を示している。
【図7】非トリプシン性のN末端切断を示す、NC4ドメインにおけるペプチドSNSDGENELCPK(配列番号12)のタンデム質量分析データ(LC−ESI−QTof MS)を示す図である。バンドをゲルから切り取り、トリプシンで消化し、その後、オンラインでESI−Qtof MSを用いた逆相クロマトグラフィーを行った。ペプチドを脱アミド化して、このペプチド配列におけるD(アスパラギン酸塩)がウシ配列gi|119901059)においてはN(アスパラギン)となるようにする。この注釈付きのスペクトルは、BioLynx(ペプチド配列決定、Waters Djupdalsvagen 12−14 Sollentuna Sweden)において生じたものである。ペプチドのデルタ質量は0.01Daであり、yイオンの平均誤差は<0.02Daであり、強度の閾値は2%に設定した。同一の実験のデータを図12cに示す。
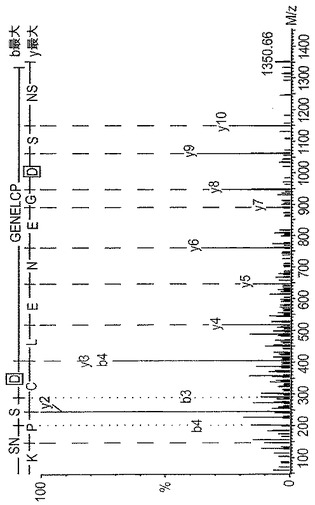
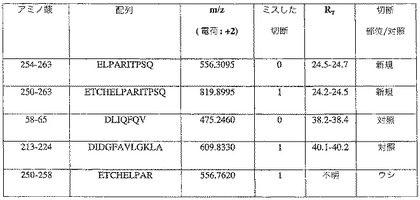
【図8】図8Aは、SDSの存在下及び不在下でRA試料及びOA試料を比較する、ネオエピトープ抗血清を用いる免疫アッセイを示すグラフである。1つのRA患者及び1つのOA患者から得た滑液を、SDSの存在下又は不在下で分析した。試料は、1:10、1:40、及び1:160に希釈した。図8Bは、4〜16%ゲル上でSDS−ポリアクリルアミド電気泳動し、その後、電気泳動でニトロセルロース膜に移し、PARITPSQ(配列番号13)切断ネオエピトープに対する抗体を用いて染色した、関節疾患を有する患者から得た滑液試料のウェスタンブロットを示すグラフである。結合したIgGを、ペルオキシダーゼ結合抗ウサギIgG抗体を用いて検出した。ネオエピトープを含む、選択滑液における20kDaの断片の存在に注目されたい。この断片は、MMP−13消化の際に関節軟骨から放出された断片よりも多少小さいものである。
【図9】示した切断部位を有するIX型コラーゲン分子の概略図である。細い線は非コラーゲンドメイン(NC)に相当し、太い線はコラーゲンドメイン(COL)に相当する。矢印は、同定され記載されている切断部位の位置を示している。
【図10】それぞれ、ヒト及びウシの軟骨におけるIX型コラーゲンの切断配列を示す図である。
【図11a】図1の2Dゲルから得たスポット5の、MALDI MS分析の結果を示す表である。Mascotペプチドマスフィンガープリントソフトウェア(Matrix Science Inc、Boston、MA、USA)を用いたデータベース検索の結果、(IX型)コラーゲンα1が同定された。NC4ドメインから得たペプチドのみがスペクトルにおけるピークに適合した。適合したペプチドを、ウシ配列gi|119901059に関する開始位置及び終了位置と共に表にする。M(expt)は、ペプチドの測定されたモノアイソトピック質量であり、Hは水素であり、「デルタ」は、適合ペプチドの、測定された質量と理論的に計算された質量との間の差を意味する。「ミス」は、適合ペプチド配列内にどれだけ多くのミスした切断が存在するかを示す。「配列」は、N末端及びC末端の両方にハイフンで分離された1つの余分なアミノ酸を有する適合ペプチドの配列を示す。「修飾」は、質量分析で得られたM(expt)にペプチド配列を適合させるために必要な、ペプチド配列における修飾を説明するものである。認められた修飾(変動的)は、メチオニンの酸化、及びヒドロキシプロリンを形成するためのプロリン残基のヒドロキシル化であった。システイン残基はリストされなかったため、全てのシステイン残基はカルバミドメチル化されたと考えられた。
【図11b】図1の2Dゲルから得たスポット8の、MALDI MS分析の結果を示す表である。その後、Mascotペプチドマスフィンガープリントソフトウェアを用いてデータベース検索を行った結果、(IX型)コラーゲンα1が同定された。NC4ドメイン及びCOL3ドメインの両方におけるペプチドがスペクトルにおけるピークに適合した。MMP−13処理した軟骨移植片から放出された50kDaの断片を分析した後(図4)、このスペクトルを、切断部位のペプチドに適合するピークの存在について調べた。それはこれらのMALDIデータにおいても存在した(観察された質量=1274.628)。適合したペプチドを、ウシ配列gi|119901059に関する開始位置及び終了位置と共に表にする。表の説明については図11aにおける記載を参照されたい。
【図11c】図1の2Dゲルから得たスポット14の、MALDI MS分析の結果を示す表である。その後、Mascotペプチドマスフィンガープリントソフトウェアを用いてデータベース検索を行った結果、(IX型)コラーゲンα1が同定された。NC4ドメイン、COL3ドメイン、及びCOL2ドメインから得たペプチドがスペクトルにおけるピークに適合した。適合したペプチドを、ウシ配列gi|119901059に関する開始位置及び終了位置と共に表にする。表の説明については図11aにおける記載を参照されたい。
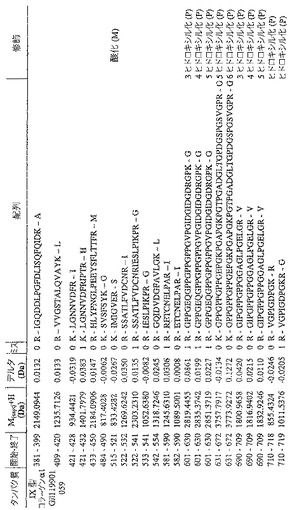
【図12a】Mascot「MSMS lons search」(Matrix Science Inc.Boston、MA、USA)を用いたデータベース検索に従った、ウシ(IX型)コラーゲンα1に適合するペプチドを示す表である。28kDaの断片を2Dゲルから切り出し、AspNでゲル内消化し、ESI−Qtof MS/MSと共にオンラインで逆相ナノLCを用いて分析した。これは、図3に示したものと同一の実験から得られたデータである。「M(expt)」は、適合ペプチドのモノアイソトピック質量を意味し、そうでなければ、この表記は図11におけるものと同一である。イオンスコアは、観察された適合がランダムヒットである可能性に基づくものである(41)。値が大きいほど良好なヒットであることを示す。適合したペプチドを、ウシ配列gi|119901059に関する開始位置及び終了位置と共に表にする。
【図12b】28kDaのウシ断片のAspN消化物において、推定される、トリプシンにより生じる部位が不在であることを示す表である。図3及び図12aと同一の実験であるが、推定される「トリプシン性」ペプチドに適合するAspN消化物においてペプチドについて検索したところ、切断部位のペプチドが明らかになった。トリプシン消化に典型的な他のペプチドは同定されなかった。適合したペプチドを、ウシ配列gi|119901059に関する開始位置及び終了位置と共に表にする。
【図12c】28kDaのバンドにおける、ウシ(IX型)コラーゲンα1に適合するペプチドを示す表である。28kDaのバンドを2Dゲルから切り出し、トリプシンでゲル内消化し、Qtofセットアップを用いて分析し、データを用いてMascot「MSMS Ions search」でNCBI nrデータベースの検索を行った。適合したペプチドを、ウシ配列gi|119901059に関する開始位置及び終了位置と共に表にする(図7)。「修飾」は、質量分析で得られたm/zにペプチド配列を適合させるために必要な、ペプチド配列における修飾を説明するものである。認められた修飾(変動的)は、メチオニンの酸化、脱アミド化(アスパラギン及びグルタミン)、並びにヒドロキシプロリンであった。システイン残基はリストされなかったため、全てのシステイン残基はカルバミドメチル化されたと考えられた。
【図13】ウシ(IX型)コラーゲンα1鎖に適合するペプチドを示す表である。50kDaの、MMP−13により生じた試料を1Dゲルから切り出し、トリプシンでゲル内消化し、逆相LC ESI−IonTrap MSセットアップを用いて分析した。データを、Mascot「MSMS Ions search」を用いるデータベース検索によりさらに分析した。これは、図4bと同一の実験から得られたデータであった。適合したペプチドを、ウシ配列gi|119901059に関する開始位置及び終了位置と共に表にする。表の説明については図12a及び12cを参照されたい。
【図14】30kDaの断片における(IX型)コラーゲンα1に適合するヒトペプチドを示す表である。16%SDS PAGEで分離したMMP−13消化培地から得た、Asp−N消化した30kDaのバンドのIonTrap分析から得られたデータベース検索の結果である。
【図15】「含有質量リスト」を示す表である。(IX型)ヒトコラーゲンα1のNC4のアミノ酸配列に由来する5つのペプチド質量を、逆相LC ESI−Qtof MSセットアップによるMSMS分析のために選択した。最初の2つは、MMP−13消化培地の30kDaのバンドのAsp−N消化物において見られた、2つの最もC末端のペプチドである。3つ目及び4つ目は、平行して実施した逆相LC ESI.IonTrap MSによって容易に検出され、対照として含まれた。最後の5つ目は、切断部位がウシにおけるものと同一である場合に予想されるC末端ペプチドに相当する。
【発明を実施するための形態】
【0021】
まずウシでの研究について考察し、さらに、ヒトにおいて本発明の詳細について記載する。
【0022】
ウシ鼻軟骨のインターロイキン−1誘導性の分解により、IX型コラーゲンα1鎖のNC4ドメインなどの断片の放出が明らかになった。NC4ドメインのN末端部分におけるエピトープに対する本発明の新規なペプチド抗体により、その所見が裏付けられた。
【0023】
2つの最も豊富な断片の質量分析により、最も小さい断片が、配列ETCNELPAR(配列番号1)(−ETCNELPAR258−COOH)及び−ITPGARSP(配列番号2)(NH2−ITPGARSP)においてアルギニン−258とイソロイシン−259との間の新規な部位で切断される、ほぼ完全なNC4ドメインを含むことが明らかになった。大きい方の断片は、NC4ドメインと、NC3ドメインに近い、COL3におけるグリシン−400とスレオニン−401との間に位置する新規な切断部位、RGPPGPPGPPGPSG−(配列番号3)(RGPP(OH)GP(OH)PGPP(OH)GPSG400−COOH)及びNH2−TIGFHDGD(配列番号4)(NH2−TIGFHDGD−)を有するCOL3ドメインとを含んでいた。
【0024】
ウシでの研究の結果
ウシのアミノ酸配列
この研究における質量分析に適合したウシ(IX型)コラーゲンα1のアミノ酸配列は、「国立生物工学情報センター」(www.ncbi.nlm.nih.gov)の非冗長(nr)データベースにおいて、gi|119901059(22−Dec−06)という名前で見ることができる。ヒト(IX型)α1配列(SwissProt P20849(http://www.expasy.ch/sprot/)との、NCBIから得たこの配列のアラインメントにより、N末端及びC末端の主な相違が明らかになる(付録1)。ウシ配列のN末端部分はヒト配列よりも332アミノ酸長い。この研究において、全ての質量分析データはこの「付加的な」配列のいずれの部分にも適合しなかった。ウシの配列は、ヒトの対応物の最初の642個のアミノ酸に対して90%の類似性を示すが、C末端における279個のアミノ酸は存在しない。このウシ配列は、置換されるべきアーチファクトであると考えられる。したがって、本発明者らは、本明細書においてはヒトの配列を参照して数字の注釈を付けることを選択した。データベース検索の結果は、それらのウシ配列での数字を維持している。
【0025】
ウシ鼻軟骨移植片の培養
IL−1で刺激した軟骨移植片から得た培地に放出されたタンパク質(3、6、9、12、及び16日)を、2D SDS PAGEを用いて分離し、ペプチドマスフィンガープリンティングを介して同定した。3日目及び6日目に得たゲルは、とりわけ最上部で不規則な分離を示し、これは、ゲル内に入るには大きすぎる、インタクトな断片化されたアグリカンを大量に含む試料に典型的である。このプロジェクトにおいて、アグリカンが放出された後、しかし主要なコラーゲン放出の前に生じる、その後の分解が焦点であった(42)。(IX型)コラーゲンα1は、12日目及び16日目から得た培地の2Dゲル上では、異なる位置で同定された(典型的なゲルを図1に示す)。IL−1の不在下でインキュベートした対照試料においては、同じスポットは存在していなかった(データは図示していない)。速く移動したタンパク質スポットの組は28kDaの見かけの質量を有しており、ペプチドマスフィンガープリンティングにより見られる配列包括度(13個のピークがペプチドに適合、図11a)により、それらが(IX型)コラーゲンα1のNC4ドメインの主要な部分に相当することが示された。約50kDaの見かけの質量で移動する(IX型)コラーゲンα1断片のペプチドマスフィガープリンティングにより、ペプチドに適合した23個のピークが明らかになり(図11b)、そのうちの10個はCOL3ドメインに由来するものであり、残りはNC4に由来するものであった。(IX型)コラーゲンα1と同定された、最も遅く移動するスポット(100kDaの見かけの質量)のペプチド包括度は、NC4及びCOL3に加えて、COL2ドメインの一部を含んでいた(25個のピークがペプチドに適合、図11c)。これは架橋部分又は全サイズの(IX型)コラーゲンα1に相当するものであり得、さらなる研究は行わなかった(未知のウシC末端配列)。
【0026】
ペプチド抗体
同定された(IX型)コラーゲンα1断片の研究を裏付けるため、またその研究をさらに可能にするために、(IX型)α1のNC4ドメインのN末端部分から得たペプチド(NH2−CGQDDLPGFDLISQFQ64−CONH2)(配列番号14)を、添加した、キーリンペットヘモシアニンに結合したシステインを用いて合成し、ウサギを免疫化するために用いた。このNC4抗血清は、固相ELISAにおいてNC4ペプチドへの結合を示した(力価1:50000)。結合はNC4ペプチドを添加することにより阻害された。0.3ng/mlのペプチド濃度は、結合の効率的な阻害を示した。
【0027】
ウェスタンブロット
ポリクローナルNC4抗血清を用いたウェスタンブロットにより、SDS PAGEで分離された、IL−1で刺激した軟骨培養物から得た培地において、28kDa及び50kDaで(IX型)コラーゲンα1断片の存在が裏付けられた。40kDaに対応する位置にさらなるバンドがあることにより、他の2つよりも多少早く最初に放出された別の(IX型)コラーゲンα1要素の存在が示された(図2)。これらの3つの断片は、それぞれ28kDa、40kDa、及び50kDaの断片であるとみなされる。
【0028】
切断部位の同定
IL−1で刺激したウシ鼻軟骨から得た培地内に放出された(IX型)コラーゲンα1断片の存在を受けて、本発明者らは切断部位を探求した。28kDaの断片を含む2Dゲルの切片をトリプシン又はAspNで処理し、その後、LC MSMS(Qtof)を行った。NCBI非冗長(nr)、カルバミドメチル(C)、変動的な酸化(M)、及び変動的なヒドロキシル化(P、K)を用いたMascotデータベース検索(Matrix Science Inc.Boston MA、USA)では、NC4ドメインからのペプチドが同定されただけであった。トリプシン消化した試料及びAspN消化した試料の両方において見られた最もC末端のペプチドはETCNELPAR258(配列番号1)であった(図3)。これは、トリプシンにより両端で正確に切断されたペプチド、及びAspNによりN末端は正確に切断されているがC末端の切断はプロセシングの前に別の酵素により媒介されなくてはならないペプチドに相当する。図2a及び2bにおいて、AspN消化された試料から得たMSMSデータに適合するペプチドを表にする。トリプシンの特異性を用いた、AspN消化物におけるペプチドについてのMascot MSMS Ion searchは、「切断」ETCNELPAR258(配列番号1)ペプチドのみを示し、このことは、同定された断片に対してトリプシンの攻撃がなかったことを示していた。28kDaのスポットのトリプシン消化物は、他のNC4ペプチドに加えて、アミノ酸Phe31−Lys46、Val33−Lys46、及びSer35−Lys46をそれぞれカバーする3つのN末端ペプチドを含んでいた(図12c及び図7)。後者の2つは共にN末端に非トリプシン切断を有しており、このことは、2つの新たなN末端が存在していることを示している。異なるN末端の存在は、コラゲナーゼ消化後に単離されたNC4においてこれまでに報告されているが(43、44)、本発明者らは自らの一連のデータにおいてこれらの切断を全く観察しなかった。
【0029】
50kDaの(IX型)コラーゲンα1断片をトリプシンでゲル内消化し、逆相ESI−IonTrap MSによって分析した。9個のペプチドを配列決定し、それらは、最もC末端のペプチド(RGPP*GPP*GPP*GPSG400)(配列番号3)が、用いたトリプシンによっては生じないC末端切断を有している(P*はヒドロキシプロリン残基を示す。データは図示していない)、(IX型)コラーゲンα1のNC4−COL3ドメインに適合した。
【0030】
主要ではない40kDaの断片を、NC4ドメインのみに配列切断を有する(IX型)コラーゲンα1として同定した。それについてはさらなる研究は行わなかった。
【0031】
ウシ鼻軟骨のMMP−13消化
MMP−13を用いてウシ鼻軟骨を24時間消化すると、IL−1でのインキュベーションと同様、軟骨に視覚的影響があり、組織が透明になり始めた。NC4抗血清を用いた培地のウェスタンブロットにより、NC4ドメインを含む2つの(IX型)コラーゲンα1断片が存在することが示された。SDS−PAGEでは、抗NC4ペプチド抗血清を用いたウェスタンブロットにより示されるように、それらは一様に28kDa及び50kDaの断片に移動した(図4a)。長時間曝露すると、IL−1処理した軟骨から得た培地において見られるものに類似した、弱い40kDaのバンドも出現した(図4a、挿入図)。クマシー染色したゲルから得た28kDa及び50kDaの2つのバンドを有するゲル切片を、それぞれAspN及びトリプシンで消化した。逆相LC ESI−IonTrap MSによる分析により、(IX型)コラーゲンα1のN末端断片を同定した。28kDaの断片において見られた最もC末端のペプチドはETCNELPAR258(配列番号1)であり、これは、12日目及び16日目の、IL−1で刺激したウシ軟骨培地試料におけるものと同じペプチドである(図3)。MMP−13により生じた50kDaの(IX型)コラーゲンα1断片について(逆相LC IonTrap MSにより)同定されたペプチドを図13にリストする。最もC末端のペプチドは、トリプシンによっては生じないC末端切断を有しており(図4b)、このことは、新たに同定された切断部位を示している。同一のペプチドが、別のCOL3ドメインペプチドに加え、IL−1処理した軟骨の試料において検出された(データは図示していない)(同一のm/z、荷電状態、及び保持時間がMMP−13試料において見られたが、MSMSは実施しなかった)。
【0032】
考察
軟骨の分解がIL−1の刺激により誘発される、関節疾患における組織破壊のこのモデルは、組織高分子の断片化及び放出の研究において頻繁に使用されている。アグリカン断片が最初に放出され、その後にCOMP、フィブロモジュリン、そして最後にコラーゲンが放出されることが示されている(45〜47)。IL−1の存在下において培養した軟骨から得た培地において、本発明者らは、2Dゲル電気泳動を用い、その後MALDI MSを行うことによって、(IX型)コラーゲンα1断片の放出を見出した。28kDaの見かけのMWを有する、2Dゲル上で検出された最も豊富な断片は、球形のNC4ドメインの大部分に相当するものであった。等電点の大きなインターバルにわたる2Dゲル上の28kDaの断片のスポットの移動は、アスパラギン残基の脱アミド化による可能性がある(48)。実際、配列決定されたペプチドの1つは脱アミド化を示すと考えられた(図7)。この現象についてのさらなる調査は行わなかった。大きい方の50kDaの断片は、NC4、及びCOL3ドメインの大部分を含むことが示された。
【0033】
IX型コラーゲンのドメインCOL1、COL2、NC1、及びNC2がII型コラーゲン原線維と強く結合している一方で、COL3及び球形のNC4は線維表面から突き出している(49)。IX型コラーゲンのこの部分は、II型原線維と他の細胞外構造要素との間のコネクターとしての役割を果たし得る。裏付けとして、IX型コラーゲンが不在であると、軟骨は、病原性抗体による自己免疫攻撃に対してより脆弱になり得(50)、マトリリン−3を保持するその能力が低下する(51)ことが最近示された。したがって、IX型コラーゲンの断片化及び組織からの機能的NC4ドメインの喪失は、軟骨の解体のプロセスにおける重要な初期の段階を反映するものである。
【0034】
上述した、(IX型)コラーゲンα1ペプチド抗体は、NC4ドメインのN末端に近い配列に相当する合成ペプチド(C)GQDDLPGFDLISQFQ64(配列番号14)に対するものであった。このペプチドを、NC4ドメインに対する抗体に対する免疫原として以前に用いたペプチドのC末端となるように選択したが(52)、それは、これらの筆者が、彼らが用いた免疫原配列内にいくつかが位置している、複数のN末端の存在を記載していたからである。それらの抗体は、とりわけ成長プレートにおけるIX型コラーゲンの変化を記載するために用いられている(53〜56)。本発明者らの質量分析データもまた、NC4ドメインのN末端での複数の切断の存在を示したが(12c及び図7)、そのいずれも、これまでに記載されているものに対応しない。これらの切断がIL−1による刺激の結果であるか、又はこれらが刺激の前に組織内に既に存在しているかは分かっていない。
【0035】
本発明者らによる新規なNC4抗血清を用いたウェスタンブロットにより、9日目、12日目、及び16日目の培地において3つの異なるIX型コラーゲン断片が存在することが明らかに示された(図2)。そのうちの2つの見かけの質量は、約28kDa及び50kDaに対応する、2Dゲルから以前に同定されたスポットに適合する。40kDaの見かけの質量を有する3つ目の断片は、9日目の培地において既に見られた。12日目のIL−1で刺激した軟骨から得た培地において見られた40kDaの断片をトリプシン消化及び質量分析すると、(IX型)コラーゲンα1のNC4ドメインが同定された。見かけの質量は、NC4ドメインよりも大きな断片を示唆しているが、質量分析のデータは限定された包括度を示し、COL3ドメインのペプチドを全く含まなかった。最も豊富な28kDaの断片を選択して、さらに広範囲な特徴付けを行った。大きい方の断片は、コラーゲンドメインと、プロリン残基のヒドロキシル化におけるそれらの不均一性と、トリプシンなどの酵素によるプロリンのN末端の切断の困難性とから、研究するにはより複雑であると予想された。
【0036】
エレクトロスプレー質量分析(LCMSMS)を用いたオンラインの逆相液体クロマトグラフィーと組み合わせた、トリプシン及びエンドプロテアーゼAspNでの消化により、COL3ドメインに対する少数のアミノ酸からなるN末端である、NC4のC末端部分における新たな切断が明らかになった。トリプシンを用いて見出された最もC末端の断片はETCNELPAR258(配列番号1)であった(l)。C末端に加えて、ペプチドをカバーする部分を探求するために、NC4を含む断片をAspNで切断した。この酵素はPAR258とそれに続くイソロイシンとの間の切断を生じさせないにもかかわらず、AspNは、トリプシンと同一の、最もC末端のペプチドETCNELPAR258を生じさせた(図3)。AspN消化物は、トリプシンによる汚染を除き、トリプシンにより生じ得るいかなる他のペプチドも含んでいなかった。別の対照として、長い方の50kDaの(IX型)コラーゲンα1断片のAspN消化は、逆相LC ESI−IonTrap MSで検出可能なETCNELPAR(配列番号1)ペプチドを生じさせず、一方、トリプシンは、アルギニン残基で実際に切断してこの断片を生じさせた(データは図示していない)。50kDaの(IX型)コラーゲンα1断片をトリプシン消化し、その後、逆相LC ESI−IonTrapMSで分析すると、16日目のIL−1から得た培地においてCOL3ドメインにおける2つのペプチドが同定された。それらのうち最もC末端のものは、C末端の非トリプシン切断(−RGPPGPPGPPGPSG400−COOH(配列番号3)、NH2−TIGFHDGD−)(配列番号4)を有しており、このことは、この切断が組織においてIL−1での刺激の間に生じたことを示している(図13及び図4b)。28kDa及び50kDaの断片は組織から同時に放出され(図2)、したがって、前述の40kDaの断片の形成と並行して切断されると考えられる。
【0037】
先の実験において(57)、軟骨をIL−1で処理すると、MMP−13が活性な形態で現れ、9日目頃からフィブロモジュリンを切断することが示された。フィブロモジュリンにおける切断部位(−PAY63とA64YG−との間)はIX型コラーゲンにおいて見られた切断部位(−PAR258 I259TP−)とは異なるが、本発明者らは、MMP−13での軟骨の消化により、IL−1で刺激した軟骨移植片の培地において本発明者らが見出した(IX型)コラーゲンα1のNC4の切断が生じ得るかどうかを試験することとした。
【0038】
ウシ鼻軟骨移植片をMMP−13で消化したところ、実際にこの酵素は(IX型)コラーゲンα1を切断した。NC4抗血清を用いるウェスタンブロットにより示される、培地における断片を含むNC4の出現は、IL−1で誘導した切断により得られたものと非常に類似していた(図4a)。IL−1による刺激で以前に観察された、(IX型)コラーゲンα1のNC4から得られた28kDa及び50kDaの2つの断片が、MMP−13消化でウシ軟骨から培地内に放出された。9日目、12日目、及び16日目にIL−1処理した軟骨において見られた40kDaのバンドは、このブロットにおいては、より長い曝露時間にすると弱いバンドとして見ることができた(図4a、挿入図)。MMP−13消化したウシ軟骨の培地における28kDaの断片を、次に、逆相LC ESI−IonTrap MSによって(IX型)コラーゲンα1のNC4ドメインであると同定した。それは、IL−1で刺激したウシ軟骨の培地において以前に見られたものと同じである、最もC末端のペプチド(ETCNELPAR258)(配列番号1)を含んでいた。
【0039】
16日目のIL−1で刺激したウシ軟骨、及びMMP−13消化したウシ軟骨の両方の培地において見られた50kDaの断片は、NC4及びCOL3ドメインに由来するペプチドを含むことが示された。本発明者らによる分析の前に、軟骨のIL−1処理及びMMP−13処理の両方で放出された50kDaの断片の予備分析により、両方のケースにおいて、COL3ドメインのC末端部分において生じた同一のC末端切断ペプチド(3つのヒドロキシル化*残基を有する、R387GPP*GPP*GPP*GPSG400−COOH(配列番号3))が明らかになった。コラゲナーゼであるMMP−13が、IL−1処理した試料においてもこの切断を誘導した可能性がある。規定された1つ目の基質部位がコラゲナーゼのための基質部位に相当し、2つ目がゼラチナーゼなどの異なる酵素のためのものであるため、IL−1で刺激した軟骨における切断を実際に生じさせた酵素はMMP−13であると考えられる。MMP−13は、これらの2つの活性を有することが知られている酵素に相当する。
【0040】
組織成熟における段階としてのIX型コラーゲンの除去は、初期の研究において論じられている(58、59)。後者は、NC4ドメインが鉱化の開始時に除去されたこと、並びに、鉱化が進むにつれ、II型コラーゲン及びIX型コラーゲンの両方がマトリックスから除去されたことを報告した。切断は同定されておらず、それが本発明において観察された切断のいずれかと同一であったか結論を出すのは不可能である。肥大軟骨細胞におけるMMP−13の存在は、in situでのハイブリダイゼーションを用いてこれまでに観察されている(60、61)。本発明者らによる本研究において報告された結果と組み合わせると、軟骨内骨形成における段階として、IX型コラーゲンはMMP−13により成長プレートの原線維から除去されることが妥当であると思われる。
【0041】
示されたデータは、モデル系における、軟骨の重要な成分の分解を解明し、MMP−13を、IX型コラーゲンにおける特異的な切断を達成する能力を有するプロテアーゼであると特定するものである(図9、10)。非三重らせんの部位及び三重らせん部分に対する部位が見出された。本発明において開示される新規な部位は、切断分子のみを認識する抗体を開発するために用いることができるものであり、分子マーカー技術による新規な診断手順の開発において用いることができた。
【0042】
ヒトにおける研究の結果
MMP−13でのヒト関節軟骨の消化及び新規な切断部位の決定
ヒト関節軟骨は、ウシ鼻軟骨とは対照的に、MMP−13消化では消化の視認可能な兆候は示さなかった。それでも、消化培地を(NC4抗血清を用いて)ウェスタンブロットで分析したところ、反応性を示した(図5a)。バンドは30kDの見かけの質量を有しており、それにより、そのバンドは先に記載されたウシ断片から区別された。ウェスタンブロットにおいて染色されたバンドに対応するバンドを、クマシー染色したゲルから切り出し、Asp−N又はGlu−Cで消化し、逆相LC ESI−IonTrap MSで分析した。両方の消化物において、IX型コラーゲンα1のNC4が同定され(図11におけるAsp−Nの結果)、また、潜在的に新規な切断部位CHELPARITPSQ263−COOH(配列番号6)NH2−TTDERGPP−(配列番号7)に相当するペプチドが同定された。新規な切断部位は、28kDの断片において見られる、−ETCNELPAR258−COOH(配列番号1)NH2−ITPGARSP−(配列番号2)であるウシ切断部位の、5個のアミノ酸からなるC末端である。ウシ切断部位で切断された断片に適合する質量は、ヒト試料においては見られなかった。
【0043】
潜在的な切断部位ペプチドの配列データを改善し、裏付けるために、ESI−Qtof質量分析計を用いた。本発明者らが有しているQtofシステムはIonTrapよりも5倍感度が低く、したがって、3つのゲルバンドをプールし、重炭酸アンモニウム緩衝液中のエンドプロテイナーゼAsp−Nで消化してアスパラギン酸(D)及びグルタミン酸(E)の切断N末端を得た。
【0044】
潜在的な切断部位ペプチドに先立つか又はそれと同時の任意のペプチド溶出に対するMSMSの実施で機器が占領されることを防ぐために、限定された数のペプチド質量に対して同定及びMSMS配列決定の実施を行うように機器を設定した(図12)。遅く溶出することが知られている2つの内部対照NC4ペプチドと、0個又は1個のミスしたAsp−N切断を有する2つの潜在的に新規な切断部位ペプチドと、ウシ軟骨における先に同定された切断部位ペプチドと同等のヒトペプチドとを選択した。対照ペプチド及び2つの潜在的に新規な切断部位ペプチドを、ESI−IonTrap質量分析を用いて検出し、ここで本発明者らは、2つの後者のペプチドのMSMSデータを裏付けるためにESI−Qtof質量分析を用いることとする。Qtof実験において、対照ペプチド及び新規なヒト切断部位から生じる2つのペプチドの両方を配列決定し、それらがIX型配列に適合することを見出した(図5bに示す1つのペプチドのMSMSデータ)。ウシ切断部位ペプチドと同等のヒトペプチドはQtof分析においては(またIonTrap分析においても)存在しなかった。
【0045】
ネオエピトープペプチド抗血清(CPA抗血清)
MMP−13で消化したヒト軟骨の培地のウェスタンブロットを、16%SDS PAGEでの消化物の分離及びネオエピトープペプチド抗血清を用いた同定により実施した。NC4に対する抗体を用いて観察されたバンドと同じ位置にあるバンドを同定した(図6)。変形性関節炎の患者から得た滑液は(関節リウマチの患者と対照的に)、アフィニティー精製したCPA抗血清を用いた阻害ELISAにおいて強い阻害を示した(図8)。この実験はまた、SDSの添加が、CPA抗血清が滑液エピトープを認識するために必須であることを示した。
【実施例】
【0046】
(実施例1)
ウシ(IX型)コラーゲンα1のNC4ペプチドの抗血清
免疫原としての可能性を示し、BLAST検索において見られた他の配列に対する相同性を示さない、IX型コラーゲンα1のNC4ドメインのN末端部分の15merのペプチド配列を、免疫化のために選択した。配列は、ヒト、マウス、ラット、ウシ、ニワトリ、イヌ、チンパンジー、及びアカゲザルにおいて同一であり、IL−1で刺激した軟骨の培地において存在した。N末端のシステインを結合のために付加し、免疫化のための配列であるNH2−CGQDDLPGFDLISQFQ64−CONH2(配列番号14)を得た。キーホールリンペットヘモシアニン(KLH)に結合したペプチドを用いてウサギを免疫化した。ペプチド合成及び抗体の産生は、Innovagen AB(Lund、Sweden)により提供されるカスタムサービスであった。ペプチド及び抗血清は、それぞれNC4ペプチド及びNC4抗血清と呼ばれる。
【0047】
(実施例2)
ネオエピトープペプチド抗血清(ヒト)
放出された(IX型)α1断片のC末端部分に相当するペプチド(C)−PARITPSQ263−COOH(配列番号13)を合成し、キーホールリンペットヘモシアニン(KLH)に結合させ、ウサギを免疫化するために用いた。アガロースに結合したペプチドを有するカラムを用いて、抗血清をアフィニティー精製した。ペプチド合成、ペプチドカラムの作製、及び抗体の産生は、Innovagen AB(Lund、Sweden)により提供されるカスタムサービスであった。
【0048】
(実施例3)
ウェスタンブロット(ウシ)−図2、4、及び5a
MMP−13で消化したウシ軟骨の培地の試料を16%ポリアクリルアミド−SDSゲル電気泳動により分離し、次に、ニトロセルロース膜(Hybond−C、Amersham Biosciences)に移した(38)。ブロッキングは、トリス緩衝生理食塩水(TBS)(pH7.4)における3%(w/v)低脂肪粉乳及び0.2%(v/v)TWEEN20(商標)を用いて、4℃で一晩実施した。乳を除去したブロッキング溶液(上述)で膜をすすぎ、2%(w/v)乳、TBS、及び0.2%Tween20内に1:1000で希釈したNC4抗血清と共に、室温で1時間インキュベートした。すすいだ後、ニトロセルロース膜を、2%乳/TBS/Tween20内に1:30000で希釈したペルオキシダーゼ結合二次抗体(AffiniPureロバ抗ウサギIgG Jackson ImmunoResearch)と共に室温で1時間インキュベートした。自作のECL試薬(0.1MのTris−HCl(pH8.5)を20ml、DMSO内の250mMのルミノールを48μl、DMSO内の40mMのp−クマル酸を48μl、及び14μlのH2O2)を用いて、ブロットを1分間活性化した。Agfa CRONEX5医療用X線フィルムを適当な時間にわたり膜に曝露し、Agfa Curix60で自動的に現像した。
【0049】
(実施例4)
ウェスタンブロット(ヒト)
MMP−13で消化したヒト軟骨の培地のウェスタンブロッティングにおいて用いる、アフィニティー精製したネオエピトープ抗血清を、1:100に希釈し、一方で、全ての他の条件は実施例4に記載したものである。
【0050】
(実施例5)
阻害ELISA
Nunc Maxisorp(no446612)を、PBS内の0.1μg/mlのGGGPARITPSQ263−COOH(配列番号15)ペプチドで、多湿の箱の中で一晩、室温で被覆した。PBS内の2mg/mlのオボアルブミンを用いてさらなる結合を遮断した。ペプチド標準(0〜10μg/ml)又は変形性関節炎の滑液(1:10、1:40、及び1:160)を、0.8%SDS、0.5%オボアルブミン、及び0.1%ブタ血清に溶解し、多湿の箱の中で一晩、室温でsterilinプレート上でプレインキュベートした。各ペプチドの濃縮及び滑液の希釈を3回繰り返した。アフィニティー精製したネオエピトープ抗血清を、4%トリトン、1%ブタ血清内に1:1000で希釈し、プレインキュベートしたペプチド標準及び滑液と混合し、その後、Nuncプレートに移した。二次抗体は、1%ブタ血清内において1:1000の、アルカリホスファターゼ(Dako−306)に結合したブタ抗ウサギ免疫グロブリンであった。1mg/mlのパラジニトロフェニルリン酸を基質として用いた(9.7%のジエタノールアミン、0.01%MgCl2×6H2O、0.02%NaN3)。405nmでの吸光度の読み取りを、基質を添加した直後に行い、次に1時間のインキュベーションの後に行った。最初の読み取り(ゼロ読み取り)を2回目の読み取りから差し引き、同様に処理したこれらのウェルから得た結果を平均化し、ペプチドの濃縮又は試料の希釈に対してプロットした。
【0051】
(実施例6)
骨ミネラル密度を用いたNC4の存在/不在を比較する研究
橈骨遠位端骨折を有する5人の閉経後の女性を研究する。患者をギプス副子で4週間治療する。前腕骨の骨ミネラル密度(BMD)を、骨折の2、4、6、及び8週間後に二重エネルギーX線吸収測定法(DEXA)で測定する。同じ時に血清試料を採取する。BMDの結果を、血清試料におけるNC4の存在又は不在と比較する。NC4の検出を実施例5に従って行う。
【0052】
骨折部位の骨ミネラル密度は、骨折の2、4、6、及び8週間後に対照四肢から得た値の28%、46%、86%、103%であった。5つのケースの全てにおいて、骨折部位での骨ミネラル密度の変化は、NC4の存在/不在との相関を示した。2、4、6週目の場合ではNC4は検出されるが、8週目の場合には不在である。NC4の存在/不在により、いつ骨折が完全に修復されるかを正確に決定することができる。結論として、NC4の量は、骨折の治癒のモニタリングにおける新規な技術として使用することができる。
【0053】
したがって、前述のものは、本発明の原則のみを例示するに過ぎないとみなされる。さらに、当業者は多くの改変及び変更を容易に行い得るため、示され記載された厳密な構成及び操作に本発明を限定することは望ましくなく、したがって、配列の機能上の重要ではない変異を含む全ての適切な改変及び均等物が本発明の範囲内に再分類され得、小さな変異は本明細書において説明した特定の配列に同等なものであるとみなされる。
【0054】

【技術分野】
【0001】
本発明は、血清及び滑液中の、IL−1が媒介するIX型コラーゲンの切断断片のレベルを決定することにより、関節疾患などの結合組織を冒す障害、とりわけ、変形性関節炎、関節リウマチ、反応性関節炎、腱断裂、骨粗しょう症、骨軟化症、骨折修復、及び動脈硬化を検出又はモニターすることを目的とした方法に関する。より詳細には、本発明は、IX型コラーゲンα1鎖のN末端部分の付近、又はより詳細にはNC4ドメインのC末端部分の近くにある新規な切断部位、及びNC3ドメインに近いCOL3ドメインにおける新規な切断部位で切断されたタンパク質断片の存在の検出又はモニタリングに関する。本発明はまた、切断により生じるネオエピトープ及びIX型コラーゲンに固有のNC4ドメインの切断N末端部分におけるエピトープに対するネオエピトープ抗体を開示するものである。このような方法の実施に有用な診断キット及び抗体も提示される。
【背景技術】
【0002】
硝子軟骨のIX型コラーゲンには、主要な構成要素としてII型コラーゲンを有する線維網の、主要ではない構成要素が存在する。IX型コラーゲン分子は、ポリペプチド鎖α1、α2、及びα3からなるヘテロ三量体である(2)。それは、中断された三重らせんを有する原線維結合コラーゲン(FACIT)に属する。それぞれの鎖は、4個の非三重らせんドメインであるNC1、2、3、及び4に囲まれている3個の三重らせん(コラーゲン性)ドメインであるCOL1、2、及び3を含む(3)(図9も参照されたい)。ドメインの数字はC末端から数える。
【0003】
電子顕微鏡法を用いて、IX型コラーゲンがII型コラーゲン原線維の表面を装飾していること、及びNC4ドメインが球形構造を形成し、それが茎状のCOL3ドメインと共にII型コラーゲン原線維から突き出していることが示されている(4)。
【0004】
IX型コラーゲンは、II型コラーゲン分子及び他のIX型コラーゲン分子の両方への結合を介して、II型コラーゲン原線維に共有結合で架橋している(5〜7)。これらの結合は、ペプチド結合を切断しない作用物質による、成熟軟骨からのIX型コラーゲンの抽出を事実上不可能にする。
【0005】
NC4ドメインは、多くの分子、例えばヘパリン及び軟骨オリゴマーマトリックスタンパク質(COMP)に対する親和性を有することが示されているが(8〜10)、COLドメインはマトリリン−3と相互作用する(11)。相互作用するCOMP、MATN−3、及びCOL9における突然変異は、稀な遺伝性の、分布が不均一な疾患である多発性骨端異形成症を有する患者において見られており、骨端の成長及び骨化に影響する(12)。
【0006】
驚くべきことに、NC4断片は現在、軟骨破壊のバイオマーカーであることが示されている。切断は、軟骨の主要な構造的成分であるアグリカンが放出されてから、主要なコラーゲンが分解されるまでに生じる。X線観察のように破壊プロセスの後期段階の結果であるコラーゲンマーカーとは対照的に、NC4断片は、「引き返しの限界点」の前の破壊をもたらすプロセスのマーカーである。アグリカン関連マーカーは、分解進行の初期段階において、また組織の通常の適応化の際に放出され、したがって、病理学的プロセスを特別に示すものではない。さらに、アグリカンマーカーは炎症に関連することが多い。
【0007】
本発明に記載されているように、NC4の切断の後、II型コラーゲンに架橋したままであるIX型コラーゲンの大部分は、さらに切断されてその後の段階において放出される可能性を有している。本明細書における発明で開示されている、放出された断片の新たなC末端に対するネオエピトープ抗体は、このような切断を検出することができた。加えて、本発明において開示されているネオエピトープ抗血清を免疫組織化学で用いて、本明細書において報告されている新規な切断が軟骨のどこで(表面−深部、細胞周囲−領域間)生じているかを決定することができた。組織内に最初に残存していた断片の新たに形成されたN末端に対する抗体は、その後の段階におけるこのような断片の放出を検出することができた。タイミングについてのさらなる情報は、col3ドメインにおける切断により生じたネオエピトープを用いて得ることができた。多くの先行する刊行物及び特許出願が、IX型コラーゲンに特異的に結合するモノクローナル抗体を記載しているが、それらのいずれも、NC4付近の切断部位に特異的なネオエピトープ抗体については述べていない。
【0008】
特許出願WO1990008195A1は、ヒトIX型コラーゲン及びそれに由来する断片に免疫学的に結合するモノクローナル抗体の産生において抗原として有用な、ヒトIX型コラーゲンのセグメントに対応するオリゴペプチドを開示している。しかし、この特許出願において述べられているセグメントはIX型コラーゲンの病理学的破壊には関与しておらず、したがって、関節炎との関係においては期待されない。
【0009】
Inst of Nutraceutical Research PTYに譲渡されたWO2004110475A1において、発明は、関節炎の症状を有する個体に対する治療及び耐性誘導のための、NC4ドメインを含む組成物に関するものである。発明者らはまた、抗関節炎活性又は抗炎症活性を有するポリペプチドを結合組織から回収するための方法を記載している。上記の特許出願のように、抗体は、切断されていないIX型コラーゲンに対するものであり、ネオエピトープ抗体ではない。さらに、出願人らは、NC4を診断のためのマーカーとしては述べていない。
【0010】
Fibrogen Incに譲渡された特許出願WO1999021011A1は、血清中のIX型コラーゲンのレベルを決定することにより自己免疫障害及び結合組織障害を検出又はモニターするための方法を明らかにしている。IX型コラーゲンに対する抗体を調製する際に、NC4ドメインのN末端部分における配列が選択された。この抗体は、切断されていないIX型コラーゲンに対するものであり、本明細書の発明におけるような、固有の切断ネオエピトープに対するものではない。
【0011】
先行文献のいずれも、NC−4ドメインの排他的な放出をもたらす、このドメインにおいて生じる特定の切断部位については記載していない。組織が特定の疾患において完全に破壊されるという事実にも関わらず、切断の特徴は推測又は予測することができない。とりわけ配列の類似性の観点から、ウシ軟骨及びヒト軟骨において分子の切断が異なることも予想されなかった(図10)。
【0012】
したがって、特定の切断部位の発見は新規であり、新たな診断発明を開拓するものである。
【発明の概要】
【発明が解決しようとする課題】
【0013】
したがって、本発明者らは、滑液、血清、尿、及びあらゆる体液などのあらゆるヒト試料におけるIX型コラーゲン破壊の分析を容易にする、同定された新規な切断部位に対する方法として、高感度のウェスタンブロット技術及び高感度の阻害ELISAを開発した。この種のアッセイは、同定された切断が、患者における軟骨の分解及び関節の劣化において出現するかどうか、並びにいつ出現するかを明らかにするために用いることができた。本明細書の発明の特異的なネオエピトープ抗体を用いることで、アッセイを、疾患の進行の診断及び治療に対する疾患の応答のモニタリングに用いることができる。例えば、骨折の治療中のスポーツマンに対して、トレーニングプログラムを再開する最適な時期を決定するためである。
【課題を解決するための手段】
【0014】
インターロイキン−1誘導性のウシ鼻軟骨の分解を用いて、16日間にわたり組織における異化事象を研究した。培地を2D電気泳動(等電点電気泳動法及びSDS PAGE)によって分画化した。ペプチドマスフィンガープリンティングによる成分の同定により、12日目及び16日目に、IX型コラーゲンα1鎖のNC4ドメインなどのタンパク質断片の放出が明らかになった。
【0015】
NC4ドメインのN末端部分におけるエピトープに対する新規なペプチド抗体は所見を裏付けるものであり、既に9日目に、断片の1つが存在することを示した。
【0016】
2つの最も豊富な断片の質量分析により、最も小さい断片が、配列内のアルギニン−258とイソロイシン−259との間で切断されてC末端ETCNELPAR−(配列番号1)及びN末端−ITPGARSP(配列番号2)(ETCNELPAR258−COOH及びNH2−ITPGARSP)を生じさせる、ほぼ完全なNC4ドメインを含むことが明らかになった。
【0017】
大きい方の断片は、NC4ドメインと、NC3ドメインに近い、C末端RGPPGPPGPPGPSG−(配列番号3)及びN末端−TIGFHDGD(配列番号4)(RGPPGPPGPPGPSG400−COOH及び−NH2−TIGFHDGD)を生じさせる、COL3におけるグリシン−400とスレオニン−401との間に位置する切断部位を有するCOL3ドメインとを含む。複数の(IX型)コラーゲンα1のN末端配列の存在は、放出された分子が、IL−1処理の前に、又はその処理によって、元のN末端に非常に近い部位で切断されていることを示している。
【0018】
マトリックスメタロプロテイナーゼ13(MMP−13)がこの期間において活性であり、フィブロモジュリンを切断することが示されている(1)。MMP−13で処理した軟骨移植片は、IL−1処理で得られるものと同じサイズ及び同じ切断部位を有する(IX型)コラーゲンα1断片を放出することが示された。示されたデータは、MMP−13を伴うと考えられる潜在的に重要な分解事象、及び最終的なII型コラーゲンの損失に先行する分解事象を説明するものである。
【0019】
ヒト軟骨におけるIX型コラーゲンの切断のさらなる研究により、NC−4ドメインにヒトIX型コラーゲンα1鎖の特異的な切断部位もあったが、これは驚くべきことにウシコラーゲンのものとは異なる部位に生じたことが明らかになった。NC−4ドメインにおける、ヒトIX型コラーゲンα1鎖のCHELPARITPSQTTDERGPP(配列番号5)におけるグルタミン−263とスレオニン−264との間の部位における切断により、C末端CHELPARITPSQ−(配列番号6)及びN末端−TTDERGPP)(配列番号7)(−CHELPARITPSQ263−COOH及びNH2−TTDERGPP)が生じる。COL3ドメイン内の部位に相当する大きい方の断片は、ウシ及びヒトの軟骨に由来するもので同じサイズであり、ウシにおけるS399がヒトRGPPGPPGPPGPRG400TIGFHDGD408(配列番号8)においてはR399に交換されているにもかかわらず、切断は同じ部位、すなわちG400とT401との間で生じる。ヒトRGPPGPPGPPGPRG400TIGFHDGD408(配列番号8)の切断の結果、2つのネオエピトープRGPPGPPGPPGPRG(配列番号9)及びTIGFHDGD408(配列番号10)が生じる。ヒトNC−4及びCOL3の新たなネオエピトープ末端の全てに対する抗体が、標準的な方法を用いて開発され、切断断片との特異的な反応性を示した。抗体を用いて、エピトープを検出するためのELISAを開発した。これらのELISAを用いて、関節疾患などの結合組織を冒す障害、とりわけ、変形性関節炎、並びに関節リウマチ、腱断裂、骨折修復、及び動脈硬化を有する患者から得た滑液におけるエピトープの存在を示した。本発明の特徴は、本発明を限定するものとは解釈されない以下の実施例を参照することにより、より明瞭に理解されよう。
【図面の簡単な説明】
【0020】
【図1】2D SDS PAGEを用いて分離した、16日目の、IL−1で刺激したウシ鼻軟骨培地から得たタンパク質を示す図である。一次元目である等電点電気泳動は3〜10のpH範囲にわたるものであり、二次元目は4〜20%の勾配ゲルを用いた。画像の左側の数字は参照マーカータンパク質の位置を示す。輪で印を付けたスポットは、(IX型)コラーゲンα1であると同定されたスポットを示す。異なるIX型コラーゲン断片は、本文中において、28kDa断片、50kDa断片、及び100kDa断片であるとみなされている。画像において、分析した特定のスポットの例に5、8、及び14と印を付けている。これらのスポットのMALDI−TOF MS(マトリックス支援レーザー脱離イオン化飛行時間型質量分析)の結果を、以下の図11a、b、及びcに示す。
【図2】IL−1で刺激した軟骨移植片の、NC4抗血清を用いたウェスタンブロットを示す図である。IL−1で刺激した軟骨培養物の6日目、9日目、12日目、及び16日目の培地を、1D SDS PAGEゲル上で分離した。28kDa、40kDa、及び50kDaの見かけの質量での陽性シグナルが明瞭に視認可能である。
【図3】28kDaのNC4断片の最もC末端のペプチドから得たタンデム質量分析のデータを示す図である。28kDaの断片をAspNでゲル内消化し、ESI−Qtof MS(エレクトロスプレーイオン化−四重極飛行時間型質量分析)を用いてオンラインで逆相LC(液体クロマトグラフィー、C18カラム)で分析した。推定されるペプチド配列はETCNELPAR(配列番号1)である。IL−1で刺激した軟骨から得た培地(16日目)をまず、2D SDS PAGEで分離した。この注釈付きのスペクトルは、BioLynxソフトウェア(ペプチド配列決定、Waters Djupdalsvagen 12−14 Sollentuna Sweden)において生じたものであった。ペプチドのデルタ質量は<0.01Daであり、yイオンの平均誤差は0.01Daであり、強度の閾値は0.75%に設定した。
【図4a】NC4抗血清を用いた、IL−1及びMMP−13で処理した軟骨の培地のウェスタンブロットを示す図である。IL−1(16日目)又はMMP−13で処理したウシ鼻軟骨から放出されたタンパク質を16%SDS PAGEによって分離した。クマシーブルーG−250で染色したゲル上の同等の位置を切り取り、ゲル内消化し、ESI−IonTrap MS(エレクトロスプレーイオン化イオントラップ質量分析)を用いてオンラインで逆相クロマトグラフィー(C18カラム)で分析した。両方の28kDaの断片が、同一の最もC末端のペプチドETCNELPAR(配列番号1)を含んでいた(図3参照)。50kDaの断片は、P*がヒドロキシプロリンである、共通の最もC末端のペプチドRGPP*GPP*GPP*GPSG(配列番号3)を共有していた(図4b参照)。右側の挿入図は、40kDaのバンドを示す、同一のブロットの過剰曝露のバージョンである。
【図4b】ウシIX型コラーゲンの50kDaの断片における最もC末端のペプチドのタンデム質量分析を示す図である。50kDaの断片をトリプシンでゲル内消化し、IonTrap MSMSを用いてオンラインで逆相クロマトグラフィーで分析すると、最もC末端のペプチド(P*がヒドロキシプロリン残基であるRGPP*GPP*GPP*GPSG(配列番号3))のタンデム質量分析のデータが見られた。MMP−13消化溶液を16%SDS PAGEで分離し、クマシーブルーG−250で染色した。この注釈付きのスペクトルは、BioTools(Bruker Daltonics Scandinavia AB、Polygonvagen 79 Taby Sweden)において生じたものであった。許容誤差は0.2Daであり、bイオン及びyイオンの平均誤差は0.06Daであり、強度の閾値は10に設定した。
【図5a】MMP−13又はIL−1で処理したヒト軟骨の、NC4抗血清を用いたウェスタンブロットを示す図である。ヒト関節及びウシ鼻軟骨をMMP−13で消化し、後者はIL−1での刺激も行った(16日目)。消化溶液及び培地を16%ポリアクリルアミド−SDSゲル上で分離した。
【図5b】ヒトNC4断片の最もC末端のペプチドのタンデム質量分析のデータを示す図である。断片をAsp−Nでゲル内消化し、LC−Qtof MSで分析した。ロイシン及びイソロイシンは正確な同一の質量を有しており、したがって、スペクトルにおけるLはL又はIに相当する。推定されるペプチド配列はETCHELPARITPSQ(配列番号11)である。MMP−13で消化した軟骨の培地を16%ポリアクリルアミド−SDSゲル上で分画化し、その後、質量分析を行った。
【図6】MMP−13で消化したヒト関節軟骨の培地の、ネオエピトープ抗血清を用いたウェスタンブロットを示す図である。ヒト関節軟骨をMMP−13で消化した。消化溶液を16%ポリアクリルアミド−SDSゲル上で分離し、電気泳動でニトロセルロース膜に移し、それを、全ての変異体を認識するNC4における内部エピトープに対するウサギ抗血清(右パネル)、及び左パネルの、PARITPSQ263に対するネオエピトープ抗体とインキュベートした。結合した抗体を、ペルオキシダーゼ結合抗ウサギIgG抗体で検出した。切断部位全体にわたる連続的な配列を含む大きい方の断片とのネオエピトープの反応性がないことは、切断断片に対する特異性を示している。
【図7】非トリプシン性のN末端切断を示す、NC4ドメインにおけるペプチドSNSDGENELCPK(配列番号12)のタンデム質量分析データ(LC−ESI−QTof MS)を示す図である。バンドをゲルから切り取り、トリプシンで消化し、その後、オンラインでESI−Qtof MSを用いた逆相クロマトグラフィーを行った。ペプチドを脱アミド化して、このペプチド配列におけるD(アスパラギン酸塩)がウシ配列gi|119901059)においてはN(アスパラギン)となるようにする。この注釈付きのスペクトルは、BioLynx(ペプチド配列決定、Waters Djupdalsvagen 12−14 Sollentuna Sweden)において生じたものである。ペプチドのデルタ質量は0.01Daであり、yイオンの平均誤差は<0.02Daであり、強度の閾値は2%に設定した。同一の実験のデータを図12cに示す。
【図8】図8Aは、SDSの存在下及び不在下でRA試料及びOA試料を比較する、ネオエピトープ抗血清を用いる免疫アッセイを示すグラフである。1つのRA患者及び1つのOA患者から得た滑液を、SDSの存在下又は不在下で分析した。試料は、1:10、1:40、及び1:160に希釈した。図8Bは、4〜16%ゲル上でSDS−ポリアクリルアミド電気泳動し、その後、電気泳動でニトロセルロース膜に移し、PARITPSQ(配列番号13)切断ネオエピトープに対する抗体を用いて染色した、関節疾患を有する患者から得た滑液試料のウェスタンブロットを示すグラフである。結合したIgGを、ペルオキシダーゼ結合抗ウサギIgG抗体を用いて検出した。ネオエピトープを含む、選択滑液における20kDaの断片の存在に注目されたい。この断片は、MMP−13消化の際に関節軟骨から放出された断片よりも多少小さいものである。
【図9】示した切断部位を有するIX型コラーゲン分子の概略図である。細い線は非コラーゲンドメイン(NC)に相当し、太い線はコラーゲンドメイン(COL)に相当する。矢印は、同定され記載されている切断部位の位置を示している。
【図10】それぞれ、ヒト及びウシの軟骨におけるIX型コラーゲンの切断配列を示す図である。
【図11a】図1の2Dゲルから得たスポット5の、MALDI MS分析の結果を示す表である。Mascotペプチドマスフィンガープリントソフトウェア(Matrix Science Inc、Boston、MA、USA)を用いたデータベース検索の結果、(IX型)コラーゲンα1が同定された。NC4ドメインから得たペプチドのみがスペクトルにおけるピークに適合した。適合したペプチドを、ウシ配列gi|119901059に関する開始位置及び終了位置と共に表にする。M(expt)は、ペプチドの測定されたモノアイソトピック質量であり、Hは水素であり、「デルタ」は、適合ペプチドの、測定された質量と理論的に計算された質量との間の差を意味する。「ミス」は、適合ペプチド配列内にどれだけ多くのミスした切断が存在するかを示す。「配列」は、N末端及びC末端の両方にハイフンで分離された1つの余分なアミノ酸を有する適合ペプチドの配列を示す。「修飾」は、質量分析で得られたM(expt)にペプチド配列を適合させるために必要な、ペプチド配列における修飾を説明するものである。認められた修飾(変動的)は、メチオニンの酸化、及びヒドロキシプロリンを形成するためのプロリン残基のヒドロキシル化であった。システイン残基はリストされなかったため、全てのシステイン残基はカルバミドメチル化されたと考えられた。
【図11b】図1の2Dゲルから得たスポット8の、MALDI MS分析の結果を示す表である。その後、Mascotペプチドマスフィンガープリントソフトウェアを用いてデータベース検索を行った結果、(IX型)コラーゲンα1が同定された。NC4ドメイン及びCOL3ドメインの両方におけるペプチドがスペクトルにおけるピークに適合した。MMP−13処理した軟骨移植片から放出された50kDaの断片を分析した後(図4)、このスペクトルを、切断部位のペプチドに適合するピークの存在について調べた。それはこれらのMALDIデータにおいても存在した(観察された質量=1274.628)。適合したペプチドを、ウシ配列gi|119901059に関する開始位置及び終了位置と共に表にする。表の説明については図11aにおける記載を参照されたい。
【図11c】図1の2Dゲルから得たスポット14の、MALDI MS分析の結果を示す表である。その後、Mascotペプチドマスフィンガープリントソフトウェアを用いてデータベース検索を行った結果、(IX型)コラーゲンα1が同定された。NC4ドメイン、COL3ドメイン、及びCOL2ドメインから得たペプチドがスペクトルにおけるピークに適合した。適合したペプチドを、ウシ配列gi|119901059に関する開始位置及び終了位置と共に表にする。表の説明については図11aにおける記載を参照されたい。
【図12a】Mascot「MSMS lons search」(Matrix Science Inc.Boston、MA、USA)を用いたデータベース検索に従った、ウシ(IX型)コラーゲンα1に適合するペプチドを示す表である。28kDaの断片を2Dゲルから切り出し、AspNでゲル内消化し、ESI−Qtof MS/MSと共にオンラインで逆相ナノLCを用いて分析した。これは、図3に示したものと同一の実験から得られたデータである。「M(expt)」は、適合ペプチドのモノアイソトピック質量を意味し、そうでなければ、この表記は図11におけるものと同一である。イオンスコアは、観察された適合がランダムヒットである可能性に基づくものである(41)。値が大きいほど良好なヒットであることを示す。適合したペプチドを、ウシ配列gi|119901059に関する開始位置及び終了位置と共に表にする。
【図12b】28kDaのウシ断片のAspN消化物において、推定される、トリプシンにより生じる部位が不在であることを示す表である。図3及び図12aと同一の実験であるが、推定される「トリプシン性」ペプチドに適合するAspN消化物においてペプチドについて検索したところ、切断部位のペプチドが明らかになった。トリプシン消化に典型的な他のペプチドは同定されなかった。適合したペプチドを、ウシ配列gi|119901059に関する開始位置及び終了位置と共に表にする。
【図12c】28kDaのバンドにおける、ウシ(IX型)コラーゲンα1に適合するペプチドを示す表である。28kDaのバンドを2Dゲルから切り出し、トリプシンでゲル内消化し、Qtofセットアップを用いて分析し、データを用いてMascot「MSMS Ions search」でNCBI nrデータベースの検索を行った。適合したペプチドを、ウシ配列gi|119901059に関する開始位置及び終了位置と共に表にする(図7)。「修飾」は、質量分析で得られたm/zにペプチド配列を適合させるために必要な、ペプチド配列における修飾を説明するものである。認められた修飾(変動的)は、メチオニンの酸化、脱アミド化(アスパラギン及びグルタミン)、並びにヒドロキシプロリンであった。システイン残基はリストされなかったため、全てのシステイン残基はカルバミドメチル化されたと考えられた。
【図13】ウシ(IX型)コラーゲンα1鎖に適合するペプチドを示す表である。50kDaの、MMP−13により生じた試料を1Dゲルから切り出し、トリプシンでゲル内消化し、逆相LC ESI−IonTrap MSセットアップを用いて分析した。データを、Mascot「MSMS Ions search」を用いるデータベース検索によりさらに分析した。これは、図4bと同一の実験から得られたデータであった。適合したペプチドを、ウシ配列gi|119901059に関する開始位置及び終了位置と共に表にする。表の説明については図12a及び12cを参照されたい。
【図14】30kDaの断片における(IX型)コラーゲンα1に適合するヒトペプチドを示す表である。16%SDS PAGEで分離したMMP−13消化培地から得た、Asp−N消化した30kDaのバンドのIonTrap分析から得られたデータベース検索の結果である。
【図15】「含有質量リスト」を示す表である。(IX型)ヒトコラーゲンα1のNC4のアミノ酸配列に由来する5つのペプチド質量を、逆相LC ESI−Qtof MSセットアップによるMSMS分析のために選択した。最初の2つは、MMP−13消化培地の30kDaのバンドのAsp−N消化物において見られた、2つの最もC末端のペプチドである。3つ目及び4つ目は、平行して実施した逆相LC ESI.IonTrap MSによって容易に検出され、対照として含まれた。最後の5つ目は、切断部位がウシにおけるものと同一である場合に予想されるC末端ペプチドに相当する。
【発明を実施するための形態】
【0021】
まずウシでの研究について考察し、さらに、ヒトにおいて本発明の詳細について記載する。
【0022】
ウシ鼻軟骨のインターロイキン−1誘導性の分解により、IX型コラーゲンα1鎖のNC4ドメインなどの断片の放出が明らかになった。NC4ドメインのN末端部分におけるエピトープに対する本発明の新規なペプチド抗体により、その所見が裏付けられた。
【0023】
2つの最も豊富な断片の質量分析により、最も小さい断片が、配列ETCNELPAR(配列番号1)(−ETCNELPAR258−COOH)及び−ITPGARSP(配列番号2)(NH2−ITPGARSP)においてアルギニン−258とイソロイシン−259との間の新規な部位で切断される、ほぼ完全なNC4ドメインを含むことが明らかになった。大きい方の断片は、NC4ドメインと、NC3ドメインに近い、COL3におけるグリシン−400とスレオニン−401との間に位置する新規な切断部位、RGPPGPPGPPGPSG−(配列番号3)(RGPP(OH)GP(OH)PGPP(OH)GPSG400−COOH)及びNH2−TIGFHDGD(配列番号4)(NH2−TIGFHDGD−)を有するCOL3ドメインとを含んでいた。
【0024】
ウシでの研究の結果
ウシのアミノ酸配列
この研究における質量分析に適合したウシ(IX型)コラーゲンα1のアミノ酸配列は、「国立生物工学情報センター」(www.ncbi.nlm.nih.gov)の非冗長(nr)データベースにおいて、gi|119901059(22−Dec−06)という名前で見ることができる。ヒト(IX型)α1配列(SwissProt P20849(http://www.expasy.ch/sprot/)との、NCBIから得たこの配列のアラインメントにより、N末端及びC末端の主な相違が明らかになる(付録1)。ウシ配列のN末端部分はヒト配列よりも332アミノ酸長い。この研究において、全ての質量分析データはこの「付加的な」配列のいずれの部分にも適合しなかった。ウシの配列は、ヒトの対応物の最初の642個のアミノ酸に対して90%の類似性を示すが、C末端における279個のアミノ酸は存在しない。このウシ配列は、置換されるべきアーチファクトであると考えられる。したがって、本発明者らは、本明細書においてはヒトの配列を参照して数字の注釈を付けることを選択した。データベース検索の結果は、それらのウシ配列での数字を維持している。
【0025】
ウシ鼻軟骨移植片の培養
IL−1で刺激した軟骨移植片から得た培地に放出されたタンパク質(3、6、9、12、及び16日)を、2D SDS PAGEを用いて分離し、ペプチドマスフィンガープリンティングを介して同定した。3日目及び6日目に得たゲルは、とりわけ最上部で不規則な分離を示し、これは、ゲル内に入るには大きすぎる、インタクトな断片化されたアグリカンを大量に含む試料に典型的である。このプロジェクトにおいて、アグリカンが放出された後、しかし主要なコラーゲン放出の前に生じる、その後の分解が焦点であった(42)。(IX型)コラーゲンα1は、12日目及び16日目から得た培地の2Dゲル上では、異なる位置で同定された(典型的なゲルを図1に示す)。IL−1の不在下でインキュベートした対照試料においては、同じスポットは存在していなかった(データは図示していない)。速く移動したタンパク質スポットの組は28kDaの見かけの質量を有しており、ペプチドマスフィンガープリンティングにより見られる配列包括度(13個のピークがペプチドに適合、図11a)により、それらが(IX型)コラーゲンα1のNC4ドメインの主要な部分に相当することが示された。約50kDaの見かけの質量で移動する(IX型)コラーゲンα1断片のペプチドマスフィガープリンティングにより、ペプチドに適合した23個のピークが明らかになり(図11b)、そのうちの10個はCOL3ドメインに由来するものであり、残りはNC4に由来するものであった。(IX型)コラーゲンα1と同定された、最も遅く移動するスポット(100kDaの見かけの質量)のペプチド包括度は、NC4及びCOL3に加えて、COL2ドメインの一部を含んでいた(25個のピークがペプチドに適合、図11c)。これは架橋部分又は全サイズの(IX型)コラーゲンα1に相当するものであり得、さらなる研究は行わなかった(未知のウシC末端配列)。
【0026】
ペプチド抗体
同定された(IX型)コラーゲンα1断片の研究を裏付けるため、またその研究をさらに可能にするために、(IX型)α1のNC4ドメインのN末端部分から得たペプチド(NH2−CGQDDLPGFDLISQFQ64−CONH2)(配列番号14)を、添加した、キーリンペットヘモシアニンに結合したシステインを用いて合成し、ウサギを免疫化するために用いた。このNC4抗血清は、固相ELISAにおいてNC4ペプチドへの結合を示した(力価1:50000)。結合はNC4ペプチドを添加することにより阻害された。0.3ng/mlのペプチド濃度は、結合の効率的な阻害を示した。
【0027】
ウェスタンブロット
ポリクローナルNC4抗血清を用いたウェスタンブロットにより、SDS PAGEで分離された、IL−1で刺激した軟骨培養物から得た培地において、28kDa及び50kDaで(IX型)コラーゲンα1断片の存在が裏付けられた。40kDaに対応する位置にさらなるバンドがあることにより、他の2つよりも多少早く最初に放出された別の(IX型)コラーゲンα1要素の存在が示された(図2)。これらの3つの断片は、それぞれ28kDa、40kDa、及び50kDaの断片であるとみなされる。
【0028】
切断部位の同定
IL−1で刺激したウシ鼻軟骨から得た培地内に放出された(IX型)コラーゲンα1断片の存在を受けて、本発明者らは切断部位を探求した。28kDaの断片を含む2Dゲルの切片をトリプシン又はAspNで処理し、その後、LC MSMS(Qtof)を行った。NCBI非冗長(nr)、カルバミドメチル(C)、変動的な酸化(M)、及び変動的なヒドロキシル化(P、K)を用いたMascotデータベース検索(Matrix Science Inc.Boston MA、USA)では、NC4ドメインからのペプチドが同定されただけであった。トリプシン消化した試料及びAspN消化した試料の両方において見られた最もC末端のペプチドはETCNELPAR258(配列番号1)であった(図3)。これは、トリプシンにより両端で正確に切断されたペプチド、及びAspNによりN末端は正確に切断されているがC末端の切断はプロセシングの前に別の酵素により媒介されなくてはならないペプチドに相当する。図2a及び2bにおいて、AspN消化された試料から得たMSMSデータに適合するペプチドを表にする。トリプシンの特異性を用いた、AspN消化物におけるペプチドについてのMascot MSMS Ion searchは、「切断」ETCNELPAR258(配列番号1)ペプチドのみを示し、このことは、同定された断片に対してトリプシンの攻撃がなかったことを示していた。28kDaのスポットのトリプシン消化物は、他のNC4ペプチドに加えて、アミノ酸Phe31−Lys46、Val33−Lys46、及びSer35−Lys46をそれぞれカバーする3つのN末端ペプチドを含んでいた(図12c及び図7)。後者の2つは共にN末端に非トリプシン切断を有しており、このことは、2つの新たなN末端が存在していることを示している。異なるN末端の存在は、コラゲナーゼ消化後に単離されたNC4においてこれまでに報告されているが(43、44)、本発明者らは自らの一連のデータにおいてこれらの切断を全く観察しなかった。
【0029】
50kDaの(IX型)コラーゲンα1断片をトリプシンでゲル内消化し、逆相ESI−IonTrap MSによって分析した。9個のペプチドを配列決定し、それらは、最もC末端のペプチド(RGPP*GPP*GPP*GPSG400)(配列番号3)が、用いたトリプシンによっては生じないC末端切断を有している(P*はヒドロキシプロリン残基を示す。データは図示していない)、(IX型)コラーゲンα1のNC4−COL3ドメインに適合した。
【0030】
主要ではない40kDaの断片を、NC4ドメインのみに配列切断を有する(IX型)コラーゲンα1として同定した。それについてはさらなる研究は行わなかった。
【0031】
ウシ鼻軟骨のMMP−13消化
MMP−13を用いてウシ鼻軟骨を24時間消化すると、IL−1でのインキュベーションと同様、軟骨に視覚的影響があり、組織が透明になり始めた。NC4抗血清を用いた培地のウェスタンブロットにより、NC4ドメインを含む2つの(IX型)コラーゲンα1断片が存在することが示された。SDS−PAGEでは、抗NC4ペプチド抗血清を用いたウェスタンブロットにより示されるように、それらは一様に28kDa及び50kDaの断片に移動した(図4a)。長時間曝露すると、IL−1処理した軟骨から得た培地において見られるものに類似した、弱い40kDaのバンドも出現した(図4a、挿入図)。クマシー染色したゲルから得た28kDa及び50kDaの2つのバンドを有するゲル切片を、それぞれAspN及びトリプシンで消化した。逆相LC ESI−IonTrap MSによる分析により、(IX型)コラーゲンα1のN末端断片を同定した。28kDaの断片において見られた最もC末端のペプチドはETCNELPAR258(配列番号1)であり、これは、12日目及び16日目の、IL−1で刺激したウシ軟骨培地試料におけるものと同じペプチドである(図3)。MMP−13により生じた50kDaの(IX型)コラーゲンα1断片について(逆相LC IonTrap MSにより)同定されたペプチドを図13にリストする。最もC末端のペプチドは、トリプシンによっては生じないC末端切断を有しており(図4b)、このことは、新たに同定された切断部位を示している。同一のペプチドが、別のCOL3ドメインペプチドに加え、IL−1処理した軟骨の試料において検出された(データは図示していない)(同一のm/z、荷電状態、及び保持時間がMMP−13試料において見られたが、MSMSは実施しなかった)。
【0032】
考察
軟骨の分解がIL−1の刺激により誘発される、関節疾患における組織破壊のこのモデルは、組織高分子の断片化及び放出の研究において頻繁に使用されている。アグリカン断片が最初に放出され、その後にCOMP、フィブロモジュリン、そして最後にコラーゲンが放出されることが示されている(45〜47)。IL−1の存在下において培養した軟骨から得た培地において、本発明者らは、2Dゲル電気泳動を用い、その後MALDI MSを行うことによって、(IX型)コラーゲンα1断片の放出を見出した。28kDaの見かけのMWを有する、2Dゲル上で検出された最も豊富な断片は、球形のNC4ドメインの大部分に相当するものであった。等電点の大きなインターバルにわたる2Dゲル上の28kDaの断片のスポットの移動は、アスパラギン残基の脱アミド化による可能性がある(48)。実際、配列決定されたペプチドの1つは脱アミド化を示すと考えられた(図7)。この現象についてのさらなる調査は行わなかった。大きい方の50kDaの断片は、NC4、及びCOL3ドメインの大部分を含むことが示された。
【0033】
IX型コラーゲンのドメインCOL1、COL2、NC1、及びNC2がII型コラーゲン原線維と強く結合している一方で、COL3及び球形のNC4は線維表面から突き出している(49)。IX型コラーゲンのこの部分は、II型原線維と他の細胞外構造要素との間のコネクターとしての役割を果たし得る。裏付けとして、IX型コラーゲンが不在であると、軟骨は、病原性抗体による自己免疫攻撃に対してより脆弱になり得(50)、マトリリン−3を保持するその能力が低下する(51)ことが最近示された。したがって、IX型コラーゲンの断片化及び組織からの機能的NC4ドメインの喪失は、軟骨の解体のプロセスにおける重要な初期の段階を反映するものである。
【0034】
上述した、(IX型)コラーゲンα1ペプチド抗体は、NC4ドメインのN末端に近い配列に相当する合成ペプチド(C)GQDDLPGFDLISQFQ64(配列番号14)に対するものであった。このペプチドを、NC4ドメインに対する抗体に対する免疫原として以前に用いたペプチドのC末端となるように選択したが(52)、それは、これらの筆者が、彼らが用いた免疫原配列内にいくつかが位置している、複数のN末端の存在を記載していたからである。それらの抗体は、とりわけ成長プレートにおけるIX型コラーゲンの変化を記載するために用いられている(53〜56)。本発明者らの質量分析データもまた、NC4ドメインのN末端での複数の切断の存在を示したが(12c及び図7)、そのいずれも、これまでに記載されているものに対応しない。これらの切断がIL−1による刺激の結果であるか、又はこれらが刺激の前に組織内に既に存在しているかは分かっていない。
【0035】
本発明者らによる新規なNC4抗血清を用いたウェスタンブロットにより、9日目、12日目、及び16日目の培地において3つの異なるIX型コラーゲン断片が存在することが明らかに示された(図2)。そのうちの2つの見かけの質量は、約28kDa及び50kDaに対応する、2Dゲルから以前に同定されたスポットに適合する。40kDaの見かけの質量を有する3つ目の断片は、9日目の培地において既に見られた。12日目のIL−1で刺激した軟骨から得た培地において見られた40kDaの断片をトリプシン消化及び質量分析すると、(IX型)コラーゲンα1のNC4ドメインが同定された。見かけの質量は、NC4ドメインよりも大きな断片を示唆しているが、質量分析のデータは限定された包括度を示し、COL3ドメインのペプチドを全く含まなかった。最も豊富な28kDaの断片を選択して、さらに広範囲な特徴付けを行った。大きい方の断片は、コラーゲンドメインと、プロリン残基のヒドロキシル化におけるそれらの不均一性と、トリプシンなどの酵素によるプロリンのN末端の切断の困難性とから、研究するにはより複雑であると予想された。
【0036】
エレクトロスプレー質量分析(LCMSMS)を用いたオンラインの逆相液体クロマトグラフィーと組み合わせた、トリプシン及びエンドプロテアーゼAspNでの消化により、COL3ドメインに対する少数のアミノ酸からなるN末端である、NC4のC末端部分における新たな切断が明らかになった。トリプシンを用いて見出された最もC末端の断片はETCNELPAR258(配列番号1)であった(l)。C末端に加えて、ペプチドをカバーする部分を探求するために、NC4を含む断片をAspNで切断した。この酵素はPAR258とそれに続くイソロイシンとの間の切断を生じさせないにもかかわらず、AspNは、トリプシンと同一の、最もC末端のペプチドETCNELPAR258を生じさせた(図3)。AspN消化物は、トリプシンによる汚染を除き、トリプシンにより生じ得るいかなる他のペプチドも含んでいなかった。別の対照として、長い方の50kDaの(IX型)コラーゲンα1断片のAspN消化は、逆相LC ESI−IonTrap MSで検出可能なETCNELPAR(配列番号1)ペプチドを生じさせず、一方、トリプシンは、アルギニン残基で実際に切断してこの断片を生じさせた(データは図示していない)。50kDaの(IX型)コラーゲンα1断片をトリプシン消化し、その後、逆相LC ESI−IonTrapMSで分析すると、16日目のIL−1から得た培地においてCOL3ドメインにおける2つのペプチドが同定された。それらのうち最もC末端のものは、C末端の非トリプシン切断(−RGPPGPPGPPGPSG400−COOH(配列番号3)、NH2−TIGFHDGD−)(配列番号4)を有しており、このことは、この切断が組織においてIL−1での刺激の間に生じたことを示している(図13及び図4b)。28kDa及び50kDaの断片は組織から同時に放出され(図2)、したがって、前述の40kDaの断片の形成と並行して切断されると考えられる。
【0037】
先の実験において(57)、軟骨をIL−1で処理すると、MMP−13が活性な形態で現れ、9日目頃からフィブロモジュリンを切断することが示された。フィブロモジュリンにおける切断部位(−PAY63とA64YG−との間)はIX型コラーゲンにおいて見られた切断部位(−PAR258 I259TP−)とは異なるが、本発明者らは、MMP−13での軟骨の消化により、IL−1で刺激した軟骨移植片の培地において本発明者らが見出した(IX型)コラーゲンα1のNC4の切断が生じ得るかどうかを試験することとした。
【0038】
ウシ鼻軟骨移植片をMMP−13で消化したところ、実際にこの酵素は(IX型)コラーゲンα1を切断した。NC4抗血清を用いるウェスタンブロットにより示される、培地における断片を含むNC4の出現は、IL−1で誘導した切断により得られたものと非常に類似していた(図4a)。IL−1による刺激で以前に観察された、(IX型)コラーゲンα1のNC4から得られた28kDa及び50kDaの2つの断片が、MMP−13消化でウシ軟骨から培地内に放出された。9日目、12日目、及び16日目にIL−1処理した軟骨において見られた40kDaのバンドは、このブロットにおいては、より長い曝露時間にすると弱いバンドとして見ることができた(図4a、挿入図)。MMP−13消化したウシ軟骨の培地における28kDaの断片を、次に、逆相LC ESI−IonTrap MSによって(IX型)コラーゲンα1のNC4ドメインであると同定した。それは、IL−1で刺激したウシ軟骨の培地において以前に見られたものと同じである、最もC末端のペプチド(ETCNELPAR258)(配列番号1)を含んでいた。
【0039】
16日目のIL−1で刺激したウシ軟骨、及びMMP−13消化したウシ軟骨の両方の培地において見られた50kDaの断片は、NC4及びCOL3ドメインに由来するペプチドを含むことが示された。本発明者らによる分析の前に、軟骨のIL−1処理及びMMP−13処理の両方で放出された50kDaの断片の予備分析により、両方のケースにおいて、COL3ドメインのC末端部分において生じた同一のC末端切断ペプチド(3つのヒドロキシル化*残基を有する、R387GPP*GPP*GPP*GPSG400−COOH(配列番号3))が明らかになった。コラゲナーゼであるMMP−13が、IL−1処理した試料においてもこの切断を誘導した可能性がある。規定された1つ目の基質部位がコラゲナーゼのための基質部位に相当し、2つ目がゼラチナーゼなどの異なる酵素のためのものであるため、IL−1で刺激した軟骨における切断を実際に生じさせた酵素はMMP−13であると考えられる。MMP−13は、これらの2つの活性を有することが知られている酵素に相当する。
【0040】
組織成熟における段階としてのIX型コラーゲンの除去は、初期の研究において論じられている(58、59)。後者は、NC4ドメインが鉱化の開始時に除去されたこと、並びに、鉱化が進むにつれ、II型コラーゲン及びIX型コラーゲンの両方がマトリックスから除去されたことを報告した。切断は同定されておらず、それが本発明において観察された切断のいずれかと同一であったか結論を出すのは不可能である。肥大軟骨細胞におけるMMP−13の存在は、in situでのハイブリダイゼーションを用いてこれまでに観察されている(60、61)。本発明者らによる本研究において報告された結果と組み合わせると、軟骨内骨形成における段階として、IX型コラーゲンはMMP−13により成長プレートの原線維から除去されることが妥当であると思われる。
【0041】
示されたデータは、モデル系における、軟骨の重要な成分の分解を解明し、MMP−13を、IX型コラーゲンにおける特異的な切断を達成する能力を有するプロテアーゼであると特定するものである(図9、10)。非三重らせんの部位及び三重らせん部分に対する部位が見出された。本発明において開示される新規な部位は、切断分子のみを認識する抗体を開発するために用いることができるものであり、分子マーカー技術による新規な診断手順の開発において用いることができた。
【0042】
ヒトにおける研究の結果
MMP−13でのヒト関節軟骨の消化及び新規な切断部位の決定
ヒト関節軟骨は、ウシ鼻軟骨とは対照的に、MMP−13消化では消化の視認可能な兆候は示さなかった。それでも、消化培地を(NC4抗血清を用いて)ウェスタンブロットで分析したところ、反応性を示した(図5a)。バンドは30kDの見かけの質量を有しており、それにより、そのバンドは先に記載されたウシ断片から区別された。ウェスタンブロットにおいて染色されたバンドに対応するバンドを、クマシー染色したゲルから切り出し、Asp−N又はGlu−Cで消化し、逆相LC ESI−IonTrap MSで分析した。両方の消化物において、IX型コラーゲンα1のNC4が同定され(図11におけるAsp−Nの結果)、また、潜在的に新規な切断部位CHELPARITPSQ263−COOH(配列番号6)NH2−TTDERGPP−(配列番号7)に相当するペプチドが同定された。新規な切断部位は、28kDの断片において見られる、−ETCNELPAR258−COOH(配列番号1)NH2−ITPGARSP−(配列番号2)であるウシ切断部位の、5個のアミノ酸からなるC末端である。ウシ切断部位で切断された断片に適合する質量は、ヒト試料においては見られなかった。
【0043】
潜在的な切断部位ペプチドの配列データを改善し、裏付けるために、ESI−Qtof質量分析計を用いた。本発明者らが有しているQtofシステムはIonTrapよりも5倍感度が低く、したがって、3つのゲルバンドをプールし、重炭酸アンモニウム緩衝液中のエンドプロテイナーゼAsp−Nで消化してアスパラギン酸(D)及びグルタミン酸(E)の切断N末端を得た。
【0044】
潜在的な切断部位ペプチドに先立つか又はそれと同時の任意のペプチド溶出に対するMSMSの実施で機器が占領されることを防ぐために、限定された数のペプチド質量に対して同定及びMSMS配列決定の実施を行うように機器を設定した(図12)。遅く溶出することが知られている2つの内部対照NC4ペプチドと、0個又は1個のミスしたAsp−N切断を有する2つの潜在的に新規な切断部位ペプチドと、ウシ軟骨における先に同定された切断部位ペプチドと同等のヒトペプチドとを選択した。対照ペプチド及び2つの潜在的に新規な切断部位ペプチドを、ESI−IonTrap質量分析を用いて検出し、ここで本発明者らは、2つの後者のペプチドのMSMSデータを裏付けるためにESI−Qtof質量分析を用いることとする。Qtof実験において、対照ペプチド及び新規なヒト切断部位から生じる2つのペプチドの両方を配列決定し、それらがIX型配列に適合することを見出した(図5bに示す1つのペプチドのMSMSデータ)。ウシ切断部位ペプチドと同等のヒトペプチドはQtof分析においては(またIonTrap分析においても)存在しなかった。
【0045】
ネオエピトープペプチド抗血清(CPA抗血清)
MMP−13で消化したヒト軟骨の培地のウェスタンブロットを、16%SDS PAGEでの消化物の分離及びネオエピトープペプチド抗血清を用いた同定により実施した。NC4に対する抗体を用いて観察されたバンドと同じ位置にあるバンドを同定した(図6)。変形性関節炎の患者から得た滑液は(関節リウマチの患者と対照的に)、アフィニティー精製したCPA抗血清を用いた阻害ELISAにおいて強い阻害を示した(図8)。この実験はまた、SDSの添加が、CPA抗血清が滑液エピトープを認識するために必須であることを示した。
【実施例】
【0046】
(実施例1)
ウシ(IX型)コラーゲンα1のNC4ペプチドの抗血清
免疫原としての可能性を示し、BLAST検索において見られた他の配列に対する相同性を示さない、IX型コラーゲンα1のNC4ドメインのN末端部分の15merのペプチド配列を、免疫化のために選択した。配列は、ヒト、マウス、ラット、ウシ、ニワトリ、イヌ、チンパンジー、及びアカゲザルにおいて同一であり、IL−1で刺激した軟骨の培地において存在した。N末端のシステインを結合のために付加し、免疫化のための配列であるNH2−CGQDDLPGFDLISQFQ64−CONH2(配列番号14)を得た。キーホールリンペットヘモシアニン(KLH)に結合したペプチドを用いてウサギを免疫化した。ペプチド合成及び抗体の産生は、Innovagen AB(Lund、Sweden)により提供されるカスタムサービスであった。ペプチド及び抗血清は、それぞれNC4ペプチド及びNC4抗血清と呼ばれる。
【0047】
(実施例2)
ネオエピトープペプチド抗血清(ヒト)
放出された(IX型)α1断片のC末端部分に相当するペプチド(C)−PARITPSQ263−COOH(配列番号13)を合成し、キーホールリンペットヘモシアニン(KLH)に結合させ、ウサギを免疫化するために用いた。アガロースに結合したペプチドを有するカラムを用いて、抗血清をアフィニティー精製した。ペプチド合成、ペプチドカラムの作製、及び抗体の産生は、Innovagen AB(Lund、Sweden)により提供されるカスタムサービスであった。
【0048】
(実施例3)
ウェスタンブロット(ウシ)−図2、4、及び5a
MMP−13で消化したウシ軟骨の培地の試料を16%ポリアクリルアミド−SDSゲル電気泳動により分離し、次に、ニトロセルロース膜(Hybond−C、Amersham Biosciences)に移した(38)。ブロッキングは、トリス緩衝生理食塩水(TBS)(pH7.4)における3%(w/v)低脂肪粉乳及び0.2%(v/v)TWEEN20(商標)を用いて、4℃で一晩実施した。乳を除去したブロッキング溶液(上述)で膜をすすぎ、2%(w/v)乳、TBS、及び0.2%Tween20内に1:1000で希釈したNC4抗血清と共に、室温で1時間インキュベートした。すすいだ後、ニトロセルロース膜を、2%乳/TBS/Tween20内に1:30000で希釈したペルオキシダーゼ結合二次抗体(AffiniPureロバ抗ウサギIgG Jackson ImmunoResearch)と共に室温で1時間インキュベートした。自作のECL試薬(0.1MのTris−HCl(pH8.5)を20ml、DMSO内の250mMのルミノールを48μl、DMSO内の40mMのp−クマル酸を48μl、及び14μlのH2O2)を用いて、ブロットを1分間活性化した。Agfa CRONEX5医療用X線フィルムを適当な時間にわたり膜に曝露し、Agfa Curix60で自動的に現像した。
【0049】
(実施例4)
ウェスタンブロット(ヒト)
MMP−13で消化したヒト軟骨の培地のウェスタンブロッティングにおいて用いる、アフィニティー精製したネオエピトープ抗血清を、1:100に希釈し、一方で、全ての他の条件は実施例4に記載したものである。
【0050】
(実施例5)
阻害ELISA
Nunc Maxisorp(no446612)を、PBS内の0.1μg/mlのGGGPARITPSQ263−COOH(配列番号15)ペプチドで、多湿の箱の中で一晩、室温で被覆した。PBS内の2mg/mlのオボアルブミンを用いてさらなる結合を遮断した。ペプチド標準(0〜10μg/ml)又は変形性関節炎の滑液(1:10、1:40、及び1:160)を、0.8%SDS、0.5%オボアルブミン、及び0.1%ブタ血清に溶解し、多湿の箱の中で一晩、室温でsterilinプレート上でプレインキュベートした。各ペプチドの濃縮及び滑液の希釈を3回繰り返した。アフィニティー精製したネオエピトープ抗血清を、4%トリトン、1%ブタ血清内に1:1000で希釈し、プレインキュベートしたペプチド標準及び滑液と混合し、その後、Nuncプレートに移した。二次抗体は、1%ブタ血清内において1:1000の、アルカリホスファターゼ(Dako−306)に結合したブタ抗ウサギ免疫グロブリンであった。1mg/mlのパラジニトロフェニルリン酸を基質として用いた(9.7%のジエタノールアミン、0.01%MgCl2×6H2O、0.02%NaN3)。405nmでの吸光度の読み取りを、基質を添加した直後に行い、次に1時間のインキュベーションの後に行った。最初の読み取り(ゼロ読み取り)を2回目の読み取りから差し引き、同様に処理したこれらのウェルから得た結果を平均化し、ペプチドの濃縮又は試料の希釈に対してプロットした。
【0051】
(実施例6)
骨ミネラル密度を用いたNC4の存在/不在を比較する研究
橈骨遠位端骨折を有する5人の閉経後の女性を研究する。患者をギプス副子で4週間治療する。前腕骨の骨ミネラル密度(BMD)を、骨折の2、4、6、及び8週間後に二重エネルギーX線吸収測定法(DEXA)で測定する。同じ時に血清試料を採取する。BMDの結果を、血清試料におけるNC4の存在又は不在と比較する。NC4の検出を実施例5に従って行う。
【0052】
骨折部位の骨ミネラル密度は、骨折の2、4、6、及び8週間後に対照四肢から得た値の28%、46%、86%、103%であった。5つのケースの全てにおいて、骨折部位での骨ミネラル密度の変化は、NC4の存在/不在との相関を示した。2、4、6週目の場合ではNC4は検出されるが、8週目の場合には不在である。NC4の存在/不在により、いつ骨折が完全に修復されるかを正確に決定することができる。結論として、NC4の量は、骨折の治癒のモニタリングにおける新規な技術として使用することができる。
【0053】
したがって、前述のものは、本発明の原則のみを例示するに過ぎないとみなされる。さらに、当業者は多くの改変及び変更を容易に行い得るため、示され記載された厳密な構成及び操作に本発明を限定することは望ましくなく、したがって、配列の機能上の重要ではない変異を含む全ての適切な改変及び均等物が本発明の範囲内に再分類され得、小さな変異は本明細書において説明した特定の配列に同等なものであるとみなされる。
【0054】
【特許請求の範囲】
【請求項1】
血液及び滑液におけるIX型コラーゲンのα−1鎖の切断タンパク質断片のレベルを決定するステップを含む、結合組織を冒すヒトの障害を検出又はモニターする方法。
【請求項2】
障害が、関節疾患、変形性関節炎、関節リウマチ、反応性関節炎、腱断裂、骨粗しょう症、骨軟化症、骨折修復、及び動脈硬化からなる群から選択される、請求項1に記載の方法。
【請求項3】
NC4が診断のためのマーカーとして用いられる、請求項1に記載の方法。
【請求項4】
断片が、IX型コラーゲンα1鎖のN末端のNC4ドメインの、このNC4ドメインのC末端部分に近い部位と、NC3ドメインに近いCOL3ドメインとでの切断により形成される、請求項1に記載の方法。
【請求項5】
決定するステップが、IX型コラーゲンの切断により生じる切断ネオエピトープとの特異的な反応性を有する抗体の使用を含む、請求項1に記載の方法。
【請求項6】
切断タンパク質断片のレベルが、ウェスタンブロット技術を用いて決定される、請求項1に記載の方法。
【請求項7】
切断タンパク質断片のレベルが、阻害ELISA分析を用いて決定される、請求項1に記載の方法。
【請求項8】
断片が、NC−4ドメインにおける、ヒトIX型コラーゲンα1鎖のCHELPARITPSQ263TTDERGPP271(配列番号5)におけるグルタミン−263とスレオニン−264との間の新規な部位での切断により生じる群から選択される、請求項1に記載の方法。
【請求項9】
断片が、COL3ドメインにおける、ヒトIX型コラーゲンα1鎖のRGPPGPPGPPGPRG400TIGFHDGD408(配列番号8)におけるグリシン−400とスレオニン−401との間の新規な部位での切断により生じる群から選択される、請求項1に記載の方法。
【請求項10】
タンパク質分解活性を伴う、軟骨における病理学的プロセスによるIX型コラーゲンの切断により生じる、NC4ドメインのC末端部分における切断ネオエピトープ。
【請求項11】
Q残基(グルタミン)が遊離していなければならず、且つ完全な必要条件である、PARITPSQ(配列番号13)を含む、請求項10に記載の切断ネオエピトープ。
【請求項12】
末端のT残基(スレオニン)が遊離していなければならず、且つ完全な必要条件である、TTDERGPP(配列番号7)を含む、請求項10に記載の切断ネオエピトープ。
【請求項13】
G残基(グリシン)が遊離していなければならず、且つ完全な必要条件である、RGPPGPPGPPGPRG(配列番号9)を含む、請求項10に記載の切断ネオエピトープ。
【請求項14】
T残基(スレオニン)が遊離していなければならず、且つ完全な必要条件である、TIGFHDGD(配列番号10)を含む、請求項10に記載の切断ネオエピトープ。
【請求項15】
PARITPSQ(配列番号13)、TTDERGPP(配列番号7)、RGPPGPPGPPGPRG(配列番号9)、及びTIGFHDGD(配列番号10)からなる群から選択される切断ネオエピトープとの特異的な反応性を有する抗体。
【請求項16】
請求項15に記載の抗体を含む、結合組織を冒すヒトの障害の検出又はモニタリングにおいて有用な診断キット。
【請求項17】
Q263が末端アミノ酸であるネオエピトープPARITPSQ263(配列番号13)の検出を含む、組織におけるコラーゲン切断の位置を決定する方法。
【請求項18】
T264が末端アミノ酸であるネオエピトープT264TDERGPP(配列番号7)の検出を含む、組織におけるコラーゲン切断の位置を決定する方法。
【請求項19】
G400が末端アミノ酸であるネオエピトープRGPPGPPGPPGPRG400(配列番号9)の検出を含む、組織におけるコラーゲン切断の位置を決定する方法。
【請求項20】
T401が末端アミノ酸であるネオエピトープT401IGFHDGD(配列番号10)の検出を含む、組織におけるコラーゲン切断の位置を決定する方法。
【請求項1】
血液及び滑液におけるIX型コラーゲンのα−1鎖の切断タンパク質断片のレベルを決定するステップを含む、結合組織を冒すヒトの障害を検出又はモニターする方法。
【請求項2】
障害が、関節疾患、変形性関節炎、関節リウマチ、反応性関節炎、腱断裂、骨粗しょう症、骨軟化症、骨折修復、及び動脈硬化からなる群から選択される、請求項1に記載の方法。
【請求項3】
NC4が診断のためのマーカーとして用いられる、請求項1に記載の方法。
【請求項4】
断片が、IX型コラーゲンα1鎖のN末端のNC4ドメインの、このNC4ドメインのC末端部分に近い部位と、NC3ドメインに近いCOL3ドメインとでの切断により形成される、請求項1に記載の方法。
【請求項5】
決定するステップが、IX型コラーゲンの切断により生じる切断ネオエピトープとの特異的な反応性を有する抗体の使用を含む、請求項1に記載の方法。
【請求項6】
切断タンパク質断片のレベルが、ウェスタンブロット技術を用いて決定される、請求項1に記載の方法。
【請求項7】
切断タンパク質断片のレベルが、阻害ELISA分析を用いて決定される、請求項1に記載の方法。
【請求項8】
断片が、NC−4ドメインにおける、ヒトIX型コラーゲンα1鎖のCHELPARITPSQ263TTDERGPP271(配列番号5)におけるグルタミン−263とスレオニン−264との間の新規な部位での切断により生じる群から選択される、請求項1に記載の方法。
【請求項9】
断片が、COL3ドメインにおける、ヒトIX型コラーゲンα1鎖のRGPPGPPGPPGPRG400TIGFHDGD408(配列番号8)におけるグリシン−400とスレオニン−401との間の新規な部位での切断により生じる群から選択される、請求項1に記載の方法。
【請求項10】
タンパク質分解活性を伴う、軟骨における病理学的プロセスによるIX型コラーゲンの切断により生じる、NC4ドメインのC末端部分における切断ネオエピトープ。
【請求項11】
Q残基(グルタミン)が遊離していなければならず、且つ完全な必要条件である、PARITPSQ(配列番号13)を含む、請求項10に記載の切断ネオエピトープ。
【請求項12】
末端のT残基(スレオニン)が遊離していなければならず、且つ完全な必要条件である、TTDERGPP(配列番号7)を含む、請求項10に記載の切断ネオエピトープ。
【請求項13】
G残基(グリシン)が遊離していなければならず、且つ完全な必要条件である、RGPPGPPGPPGPRG(配列番号9)を含む、請求項10に記載の切断ネオエピトープ。
【請求項14】
T残基(スレオニン)が遊離していなければならず、且つ完全な必要条件である、TIGFHDGD(配列番号10)を含む、請求項10に記載の切断ネオエピトープ。
【請求項15】
PARITPSQ(配列番号13)、TTDERGPP(配列番号7)、RGPPGPPGPPGPRG(配列番号9)、及びTIGFHDGD(配列番号10)からなる群から選択される切断ネオエピトープとの特異的な反応性を有する抗体。
【請求項16】
請求項15に記載の抗体を含む、結合組織を冒すヒトの障害の検出又はモニタリングにおいて有用な診断キット。
【請求項17】
Q263が末端アミノ酸であるネオエピトープPARITPSQ263(配列番号13)の検出を含む、組織におけるコラーゲン切断の位置を決定する方法。
【請求項18】
T264が末端アミノ酸であるネオエピトープT264TDERGPP(配列番号7)の検出を含む、組織におけるコラーゲン切断の位置を決定する方法。
【請求項19】
G400が末端アミノ酸であるネオエピトープRGPPGPPGPPGPRG400(配列番号9)の検出を含む、組織におけるコラーゲン切断の位置を決定する方法。
【請求項20】
T401が末端アミノ酸であるネオエピトープT401IGFHDGD(配列番号10)の検出を含む、組織におけるコラーゲン切断の位置を決定する方法。
【図1】

【図2】
【図3】
【図4a)】
【図4b)】
【図5a】
【図5b】
【図6】

【図7】
【図8】
【図9】
【図10】

【図11a】

【図11b】
【図12a】

【図12b】

【図12c】

【図13】

【図14】

【図15】
【図2】
【図3】
【図4a)】
【図4b)】
【図5a】
【図5b】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11a】
【図11b】
【図12a】
【図12b】
【図12c】
【図13】
【図14】
【図15】
【公表番号】特表2010−520449(P2010−520449A)
【公表日】平成22年6月10日(2010.6.10)
【国際特許分類】
【出願番号】特願2009−551654(P2009−551654)
【出願日】平成20年2月29日(2008.2.29)
【国際出願番号】PCT/SE2008/050227
【国際公開番号】WO2008/108723
【国際公開日】平成20年9月12日(2008.9.12)
【出願人】(509245751)アナマル メディカル アクチボラゲット (2)
【Fターム(参考)】
【公表日】平成22年6月10日(2010.6.10)
【国際特許分類】
【出願日】平成20年2月29日(2008.2.29)
【国際出願番号】PCT/SE2008/050227
【国際公開番号】WO2008/108723
【国際公開日】平成20年9月12日(2008.9.12)
【出願人】(509245751)アナマル メディカル アクチボラゲット (2)
【Fターム(参考)】
[ Back to top ]