
新規なメチル化テアフラビン及びそれを含む組成物
【課題】1又は2以上の水酸基がメチル化されたガロイル基を有し、安定性や機能性に優れた新規なメチル化テアフラビンの提供。
【解決手段】1又は2以上の水酸基がメチル化されたガロイル基を有するメチル化テアフラビンであって、下記一般式(I)〜(III)のいずれか1の式で表されることを特徴とする化合物。

【解決手段】1又は2以上の水酸基がメチル化されたガロイル基を有するメチル化テアフラビンであって、下記一般式(I)〜(III)のいずれか1の式で表されることを特徴とする化合物。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、1又は2以上の水酸基がメチル化されたガロイル基を有する新規なメチル化テアフラビンに関するものである。
【背景技術】
【0002】
植物の果実や葉等に主に含まれている成分の1つに、ポリフェノールがある。ポリフェノールとしては、例えば、フラボン類、フラボノール類、フラバノン類、イソフラボン類、アントシアニン類、フラバノール類、エラジタンニン類、フェニルプロパノイド類、アントシアニジン類、プロアントシアニジン類、カルコン類、オーロン類、フェニルエタノイド類等が挙げられる。より具体的には、没食子酸、エラグ酸、ヒドロキシチロソール、エピガロカテキン−3−O−ガレート(EGCG)、エピカテキン−3−O−ガレート(ECG)、カテキン−3−O−ガレート(CG)、ガロカテキン−3−O−ガレート(GCG)、エピカテキン(EC)、カテキン(C)、エピガロカテキン(EGC)、ガロカテキン(GC)、クロマニン、デルフィニジン、デルフィニジン3−O−グルコシド、プロシアニジンB2(PB2)、カフェ酸、クロロゲン酸、ロスマリン酸、カフェ酸フェネチルエステル、ストリクチニン、ケルセチン、イソクエルシトリン、ルチン、ミリセチン、ブテイン、スルフレチン、ルテオリン、エリオジクチオール等が挙げられる。これらのポリフェノールは、その構造に複数のフェノール性水酸基を持つことを特徴とし、抗酸化活性、血圧上昇抑制、血糖値上昇抑制、内脂肪蓄積抑制、動脈硬化抑制、抗炎症作用、抗がん性、アレルギー抑制、抗う蝕作用、消臭作用、抗菌活性等の様々な機能性が研究、報告されている。また、これらのポリフェノール以外にも、植物由来の成分に関して、様々な機能性が研究、報告されている。
【0003】
これらのポリフェノールの中でも、水酸基にメチル基が修飾されたメチル化ポリフェノールは、メチル基で修飾されていないポリフェノールに比べて、吸収性、代謝安定性、機能性等に優れているという報告が多くなされている。例えば、茶品種“青心大ぱん”、“べにほまれ”、“べにふじ”、“べにふうき”等には、エピガロカテキン−3−O−(3−O−メチル)ガレート(EGCG3”Me)やエピガロカテキン−3−O−(4−O−メチル)ガレート(EGCG4”Me)が含まれているが、これらのメチル化カテキンをマウスへ経口投与した場合の、投与60分後の血中メチル化カテキン濃度は、エピガロカテキン−3−O−ガレート(EGCG)と比較して、遊離体で約9倍高い値を示すことが報告されている(例えば、非特許文献1参照。)。また、7−ヒドロキシフラボン、7,4’−ジヒドロキシフラボン、クリシン、アピゲニンとこれらのメチル化体を比較したところ、メチル化体のほうが、Caco−2細胞を用いたヒト腸管吸収モデル試験において高い吸収性を示し、ヒト肝組織由来S9フラクションを用いた代謝安定性試験において良好な結果を示すことも報告されている(例えば、非特許文献2参照。)。さらに、マウスマスト細胞を用いたヒスタミン遊離抑制試験に関しては、EGCG3”Me、エピガロカテキン−3−O−(3,5−O−ジメチル)ガレート、及びエピ(3−O−メチル)ガロカテキン−3−O−(3,5−O−ジメチル)ガレートは、EGCGと比較して高い抗アレルギー作用を示し、かつメチル基の増加に伴ってその作用は強まる傾向を示したことも報告されている(例えば、特許文献1参照。)。
【0004】
紅茶ポリフェノールの一種であるテアフラビンにも、脂肪吸収抑制作用、抗酸化作用、血糖値上昇抑制作用や抗腫瘍作用等の様々な機能性が報告されている。これらの機能を生体内で効果的に発揮させるためには、十分量のテアフラビンを摂取することが重要だと考えられる。しかしながら、紅茶中のテアフラビン含量は少なく、かつ、酸化によって分解されやすい。このため、紅茶をそのまま飲食したとしても、テアフラビンを大量かつ安定的に摂取することは難しい。
【0005】
カテキンと同様にメチル化することにより、テアフラビンの代謝安定性や機能性を改善し得ると考えられる。実際に、紅茶においても、メチル化ポリフェノールの一種であるメチル化テアフラビンが含まれていることが報告されているが、その含量は乾燥茶葉中に約0.05%と通常のテアフラビンよりもさらに少ない(例えば、非特許文献3参照。)。
このため、メチル化テアフラビンを紅茶から大量に得ることは困難である。
【0006】
メチル化テアフラビンを人為的に得る方法としては、エピガロカテキン3−O−(3−O−メチル)ガレート等のメチル化カテキン、エピカテキン、及びエピカテキンガレートを原料として、酸化酵素により重合させる方法がある(例えば、非特許文献3参照。)。
しかしながら、天然に希少なメチル化カテキンを原料として使用するこのような方法では、そもそも原料を得ること自体が難しいことに加えて、合成されるメチル化テアフラビンは、原料となるメチル化カテキンの種類に依存する。このため、現在までに、テアフラビン3−O−(3−O−メチル)ガレート、テアフラビン3−O−(3−O−メチル)ガレート,3’−O−ガレートしか得られていない。また、非特許文献3においては、メチル化テアフラビンの機能性は明らかにされていない。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特開2008−189628号公報
【非特許文献】
【0008】
【非特許文献1】サノ(Sano)、外3名、フレグランス・ジャーナル(FRAGRANCE JOURNAL)、2000年、4号、第46〜52ページ。
【非特許文献2】ウェン(Wen)、外1名、ドラッグ・メタボリズム・アンド・ディスポジション(Drug Metabolism and Disposition)、2006年、第34巻第10号、第1786〜1792ページ。
【非特許文献3】ニシムラ(Nishimura)、外5名、ジャーナル・オブ・アグリカルチュラル・アンド・フード・ケミストリー(Journal of Agricultural and Food Chemistry) 2007年、第55巻、第7252〜7257ページ。
【発明の概要】
【発明が解決しようとする課題】
【0009】
テアフラビン等のポリフェノール類は、非常に多くの機能を有しているが、これらの機能は、ポリフェノールの種類によって異なる場合がある。同様に、メチル化ポリフェノールにおいても、ポリフェノール中のいずれの水酸基をメチル化するかによって、その作用効果や活性強度が異なる可能性が高い。このため、様々なテアフラビン類をメチル化することにより、より代謝安定性や機能性に優れたメチル化テアフラビンを得ることができると期待される。
【0010】
本発明は、上記課題を鑑みてなされたものであって、1又は2以上の水酸基がメチル化されたガロイル基を有し、安定性や機能性に優れた新規なメチル化テアフラビンを提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明者らは、上記課題を解決すべく鋭意研究した結果、テアフラビンをはじめとするポリフェノールの水酸基を効率よくメチル化することができる新規なメチルトランスフェラーゼを得、当該酵素を用いてテアフラビンを基質とした酵素反応法を行なうことにより、新規なメチル化テアフラビンを合成することにより、本発明を完成させた。
【0012】
すなわち、本発明は、
(1) 1又は2以上の水酸基がメチル化されたガロイル基を有するメチル化テアフラビンであって、下記一般式(I)〜(III)のいずれか1の式で表されることを特徴とする化合物、
【化1】
[式(I)中、R1〜R3は水素原子又はメチル基を表し、かつ、R1〜R3の少なくとも2以上がメチル基である。]
【化2】
[式(II)中、R4〜R6は水素原子又はメチル基を表し、かつ、R4〜R6の少なくとも1以上がメチル基である。]
【化3】
[式(III)中、R7〜R12は水素原子又はメチル基を表し、かつ、R7〜R12の少なくとも1以上がメチル基である。但し、R7のみがメチル基である場合を除く。]
【0013】
(2) 下記式(I)−1、(II)−1〜4、及び(III)−1〜3からなる群より選択される1の式で表されることを特徴とする、前記(1)に記載の化合物、
【化4】
【化5】
【化6】
【0014】
(3) 前記(1)又は(2)に記載の化合物を含むことを特徴とする組成物、
(4) 前記(1)又は(2)に記載の化合物を含むことを特徴とする脂肪蓄積抑制剤、(5) 前記化合物が前記式(I)−1で表される化合物であることを特徴とする前記(4)記載の脂肪蓄積抑制剤、
(6) 前記(1)又は(2)に記載の化合物を含むことを特徴とする抗酸化剤、
(7) 前記化合物が、前記式(II)−1、(II)−3、(III)−1、(III)−2、及び(III)−3で表される化合物から選択される1種以上であることを特徴とする前記(6)記載の抗酸化剤、
(8) 前記(1)又は(2)に記載の化合物を含むことを特徴とする飲食品、
(9) 前記(1)又は(2)に記載の化合物を含むことを特徴とする医薬品又は医薬部外品、
(10) 前記(1)又は(2)に記載の化合物を含むことを特徴とする化粧品、
(11) 下記式(I)−0で表される化合物、下記式(II)−0で表される化合物、及び下記式(III)−0で表される化合物からなる群より選択される1種以上の化合物を基質として、メチルトランスフェラーゼ酵素を反応させることにより、前記一般式(I)で表される化合物、前記一般式(II)で表される化合物、及び前記一般式(III)で表される化合物からなる群より選択される1種以上の化合物を合成することを特徴とする、メチル化テアフラビンの製造方法、
【化7】
(12) 前記メチルトランスフェラーゼ酵素が、エノキタケ(Flammulinavelutipes)由来の酵素であることを特徴とする前記(11)記載のメチル化テアフラビンの製造方法、
を提供するものである。
【発明の効果】
【0015】
本発明のメチル化テアフラビンは、非メチル化体であるテアフラビンと同様の生理機能を有する上に、非常に良好なpH安定性及び代謝安定性を有する。このため、本発明のメチル化テアフラビンを飲食品、医薬品、医薬部外品、化粧品等の様々な商品へ応用することにより、非メチル化体を使用するよりも、機能性や経済性に優れた製品の開発が可能となる。
【図面の簡単な説明】
【0016】
【図1】エノキタケ由来メチルトランスフェラーゼ遺伝子の塩基配列及び対応するアミノ酸配列を示した図である。
【図2】図2(A)は、配列番号2で表されるアミノ酸配列と、上記2種類の担子菌の遺伝子と、相同性検察の結果相同性が高かった上位4遺伝子とのアミノ酸配列を比較した結果である。図2(B)は、図2(A)中の1〜4に示す領域から、O−メチルトランスフェラーゼ酵素のコンセンサス配列であると推定された配列を表す図である。
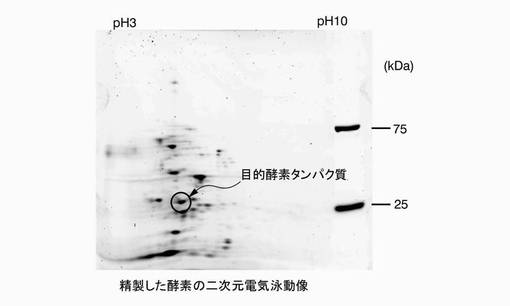
【図3】参考例1において、エノキタケ菌糸培養物から分画されたメチルトランスフェラーゼ酵素活性を有する活性画分の二次元電気泳動像である。
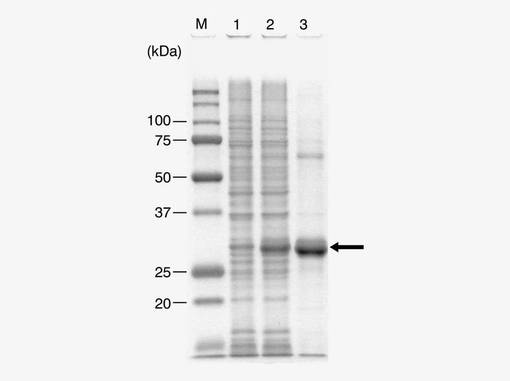
【図4】参考例2において、IPTG処理前の大腸菌のライセート(レーン1)、IPTG処理後の大腸菌のライセート(レーン2)、IPTG処理後の大腸菌のライセートからヒスチジンタグを用いて得られた精製物(レーン3)のSDS−PAGE電気泳動像である。
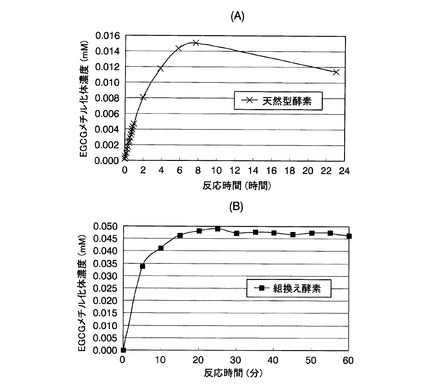
【図5】参考例5において、各反応時間後に分取した酵素反応液中のEGCGメチル化体濃度を示した図である。図5(A)が天然型酵素の結果であり、図5(B)が組換え酵素の結果である。
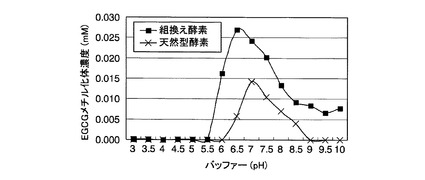
【図6】参考例6において、各pHの酵素反応液中のEGCGメチル化体濃度を示した図である。
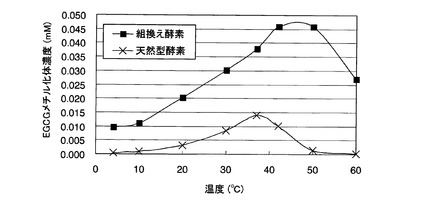
【図7】参考例7において、各温度の酵素反応液中のEGCGメチル化体濃度を示した図である。
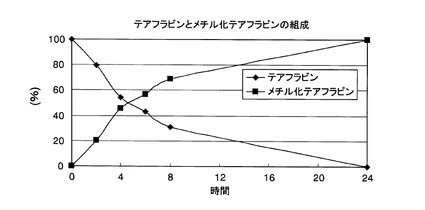
【図8】実施例2において、メチル化テアフラビン含有紅茶ポリフェノールの製造過程におけるテアフラビンとメチル化テアフラビンの組成を経時的に測定した結果を示した図である。
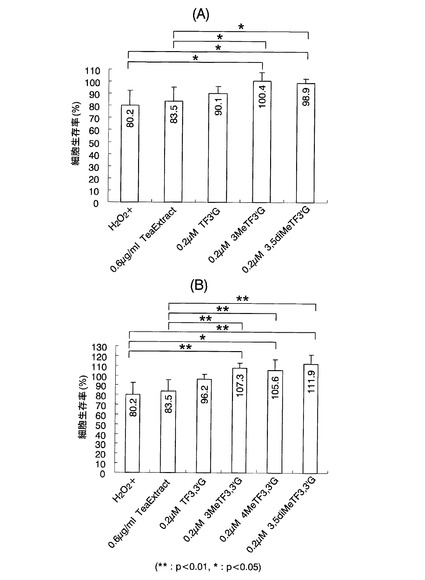
【図9】実施例3において、各種テアフラビン及びメチル化テアフラビンの残存率を反応開始2時間後に測定した結果を示した図である。図9(A)がpH5.0の結果であり、図9(B)がpH7.5の結果である。
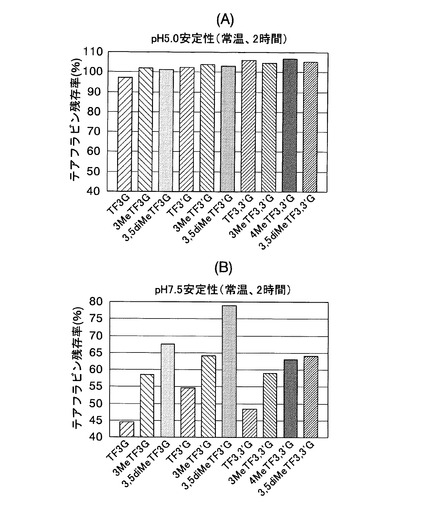
【図10】実施例4において、各種テアフラビン及びメチル化テアフラビンの残存率を反応開始60分後に測定した結果を示した図である。
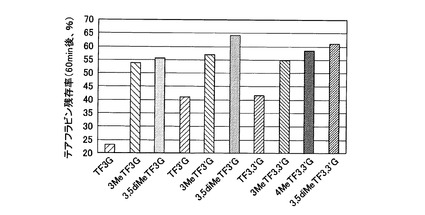
【図11】実施例5において、各処理を施した細胞群の脂肪蓄積量の測定結果を示した図である。図11(A)は各被検素材を1μMとなるように培地中に添加した場合の結果であり、図11(B)は3μMとなるように培地中に添加した場合の結果である。
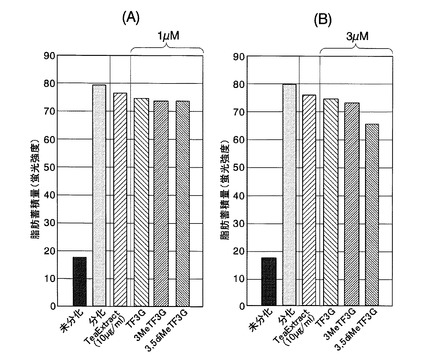
【図12】実施例6において、HepG2細胞生存率の測定結果を示した図である。図12(A)がTF3’G及びそのメチル化体を添加した場合の測定結果であり、図12(B)がTF3,3’G及びそのメチル化体を添加した場合の測定結果である。
【発明を実施するための形態】
【0017】
本発明及び本願明細書において使用する各化合物の略称を以下に示す。
TF:テアフラビン
TF3G:テアフラビン3−O−ガレート
3MeTF3G:テアフラビン3−O−(3−O−メチル)ガレート
3,5diMeTF3G:テアフラビン3−O−(3,5−O−ジメチル)ガレート
TF3’G:テアフラビン3’−O−ガレート
3MeTF3’G:テアフラビン3’−O−(3−O−メチル)ガレート
4MeTF3’G:テアフラビン3’−O−(4−O−メチル)ガレート
3,5diMeTF3’G:テアフラビン3’−O−(3,5−O−ジメチル)ガレート3,4diMeTF3’G:テアフラビン3’−O−(3,4−O−ジメチル)ガレートTF3,3’G:テアフラビン3,3’ −ジ−O−ガレート
3MeTF3,3’G:テアフラビン3−O−ガレート,3’−O−(3−O−メチル)ガレート
4MeTF3,3’G:テアフラビン3−O−ガレート,3’−O−(4−O−メチル)ガレート
3,5diMeTF3,3’G:テアフラビン3−O−ガレート,3’−O−(3,5−O−ジメチル)ガレート
Tea extract:紅茶抽出物、紅茶エキス
【0018】
本発明及び本願明細書において、テアフラビンとは、下記式(I)−0で表される化合物(TF3G)、下記式(II)−0で表される化合物(TF3’G)、又は下記式(III)−0で表される化合物(TF3,3’G)を示す。なお、これらの式で表される化合物は複数の異性体を有しているが、これらの異性体も、テアフラビンに含まれる。
また、本発明及び本願明細書において、メチル化テアフラビンとは、テアフラビンのガロイル基中の少なくとも1の水酸基がメチル化されている化合物を意味する。
【0019】
【化8】
【0020】
<新規メチルトランスフェラーゼ>
本発明者らは、新規なメチル化テアフラビンを合成するために、後記参考例1等に示すように、エノキタケ(Flammulina velutipes)由来の新規なメチルトランスフェラーゼを単離・同定した。
得られたエノキタケ由来メチルトランスフェラーゼをコードする遺伝子の塩基配列(配列番号1)及び対応するアミノ酸配列(配列番号2)を図1に示す。配列番号1で表される塩基配列の45位〜734位がコーディング領域である。
【0021】
なお、配列番号1で表される塩基配列及び配列番号2で表されるアミノ酸配列に対して、公開されているNCBI(National Center for Biotechnology Information)のデータベースを用いてホモロジー検索を行った結果、最も相同性の高い遺伝子はオオキツネタケ由来遺伝子S238N−H82(アクセッション番号:XM_001878803)であり、塩基配列で63.6%、アミノ酸配列で61.6%程度の相同性であった。
【0022】
また、全ゲノム配列が判明している担子菌類であるモデルキノコのデータベース(http://genome.jgi−psf.org/Phchr1/Phchr1.home.html)を用いて、同様にホモロジー検索を行った結果、最も相同性の高い遺伝子はO-メチルトランスフェラーゼ遺伝子(Protein ID:127115)であり、塩基配列で63.9%、アミノ酸配列で56.8%程度の相同性であった。
【0023】
図2(A)は、配列番号2で表されるアミノ酸配列と、上記2種類の担子菌の遺伝子と、相同性検察の結果相同性が高かった上位4遺伝子とのアミノ酸配列を比較した結果である。図2(A)中、「F.velutipes」は配列番号2で表されるアミノ酸配列を表す。また、「L.bicolor S238N−H82」はオオキツネタケ由来遺伝子S238N−H82、「P.chrysosporium」はモデルキノコ由来O−メチルトランスフェラーゼ遺伝子、「B.indeica subsp.」はBeijerinckia indica subsp.由来O−メチルトランスフェラーゼ ファミリータンパク質遺伝子(アクセッション番号:YP_001832797)、「B.ambifaria AMMD」はBurkholderia ambifaria AMMD由来O−メチルトランスフェラーゼ ファミリータンパク質遺伝子(アクセッション番号:YP_777310)、「B.thailandensis」はBurkholderia thailandensis MSMB43由来O−メチルトランスフェラーゼ ファミリータンパク質遺伝子(アクセッション番号:ZP_02466609)、及び「E.sakazakii」はEnterobacter sakazakii ATCC BAA−894由来遺伝子(アクセッション番号:YP_001437785)の推定アミノ酸配列をそれぞれ表す。
【0024】
本発明者らは、このアラインメントから、図2(A)中、1〜4に示す領域が、全7種類の遺伝子において保存性の高い領域であることを見出した。なお、配列番号2で表されるアミノ酸配列中、領域1は70〜88番目の領域であり、領域2は139〜148番目の領域であり、領域3は171〜175番目の領域であり、領域4は215〜223番目の領域である。さらに本発明者らは、これらの4領域のアミノ酸配列から、O−メチルトランスフェラーゼ酵素のコンセンサス配列を推定した。図2(B)は、図2(A)中の1〜4に示す領域から、O−メチルトランスフェラーゼ酵素のコンセンサス配列であると推定された配列を表す。なお、図2(B)中、「x」はいずれのアミノ酸残基であってもよいことを意味し、「(/)」は、括弧内のいずれかのアミノ酸残基であることを意味する。
【0025】
さらに、配列番号2で表されるアミノ酸配列に対して、NCBIデータベースを用いて植物由来のメチルトランスフェラーゼ酵素とのホモロジー検索を行った結果、ダイズ(Zea mays)由来遺伝子(アクセッション番号:AY279004)と、塩基配列で53.9%、アミノ酸配列で30.4%の相同性であった。同様に、イネ(Olyzasativa)由来遺伝子(アクセッション番号:AY279004)と50.7%(塩基配列)及び30.0%(アミノ酸配列)、シロイヌナズナ(Arabidopsisthaliana)由来遺伝子(アクセッション番号:MN_116065)52.3%(塩基配列)及び30.8%(アミノ酸配列)程度の相同性であった。さらに、特許文献4に記載の、茶から単離されたEGCGをメチル化可能なメチルトランスフェラーゼ酵素と比較した結果、塩基配列で53.9%、アミノ酸配列では30.4%程度の相同性であった。
【0026】
本発明者らは、テアフラビンを基質とし、同定したエノキタケ由来メチルトランスフェラーゼによる酵素反応によって、メチル化テアフラビンを得た。この結果、公知のメチル化テアフラビンである3MeTF3G以外にも、多くの新規なメチル化テアフラビンを得た。
【0027】
すなわち、本発明のメチル化テアフラビンは、下記一般式(I)〜(III)のいずれか1の式で表されることを特徴とする。
【0028】
【化9】
[式(I)中、R1〜R3は水素原子又はメチル基を表し、かつ、R1〜R3の少なくとも2以上がメチル基である。]
【0029】
【化10】
[式(II)中、R4〜R6は水素原子又はメチル基を表し、かつ、R4〜R6の少なくとも1以上がメチル基である。]
【0030】
【化11】
[式(III)中、R7〜R12は水素原子又はメチル基を表し、かつ、R7〜R12の少なくとも1以上がメチル基である。但し、R7のみがメチル基である場合を除く。]
【0031】
一般式(I)中、R1〜R3は水素原子又はメチル基を表し、かつ、R1〜R3の少なくとも2以上がメチル基である。
一般式(II)中、R4〜R6は水素原子又はメチル基を表し、かつ、R4〜R6の少なくとも1以上がメチル基である。
一般式(III)中、R7〜R12は水素原子又はメチル基を表し、かつ、R7〜R12の少なくとも1以上がメチル基である。但し、R7のみがメチル基である場合を除く。
なお、これらの一般式で表される化合物は複数の異性体を有しているが、これらの一般式で表される化合物である限り、いずれの異性体も、本発明のメチル化テアフラビンに含まれる。
【0032】
本発明のメチル化テアフラビンとしては、特に、式(I)−1で表される化合物(3,5diMeTF3G)、式(II)−1で表される化合物(3MeTF3’G)、式(II)−2で表される化合物(4MeTF3’G)、式(II)−3で表される化合物(3,5diMeTF3’G)、式(II)−4で表される化合物(3,4diMeTF3’G)、式(III)−1で表される化合物(3MeTF3,3’G)、式(III)−2で表される化合物(4MeTF3,3’G)、及び式(III)−3で表される化合物(3,5diMeTF3,3’G)であることが好ましい。
【0033】
【化12】
【0034】
【化13】
【0035】
【化14】
【0036】
本発明のメチル化テアフラビンは、エノキタケ由来メチルトランスフェラーゼを用いた酵素反応により初めて合成されたものである。紅茶にはテアフラビンが含まれており、かつ生茶葉由来のメチルトランスフェラーゼも公知である(例えば、特開2007−306806号公報)。にもかかわらず、今までこれらのメチル化テアフラビンが得られていなかった理由は明らかではないが、生茶葉由来メチルトランスフェラーゼがカテキン類に対する特異性が高く、テアフラビン類に対する特異性が十分ではない可能性や、エノキタケ由来メチルトランスフェラーゼの酵素活性が、生茶葉由来メチルトランスフェラーゼ等の公知のメチルトランスフェラーゼよりも高い可能性がある。
【0037】
本発明のメチル化テアフラビンは、テアフラビンをメチルトランスフェラーゼによりメチル化することによって製造することができる。より具体的には、例えば、1種類又は2種類以上のメチルトランスフェラーゼを、pH5〜12、好ましくはpH6〜9、より好ましくはpH6.5〜8.5の緩衝液に懸濁し、さらにテアフラビン含有物とメチル基の供与体とを加えて、0〜80℃、好ましくは10〜50℃、より好ましくは30〜42℃で反応させることにより、酵素反応液中にメチル化体を得ることができる。なお、テアフラビン含有物としては、紅茶、紅茶抽出物、紅茶ポリフェノール、テアフラビン標準品等を用いることができる。また、メチル基の供与体としては、S−adenosyl−L−methionine (SAM)等の、一般的にフェノール性水酸基のメチル化の際に用いられる物質の中から、適宜選択して用いることができる。
【0038】
本発明のメチル化テアフラビンを製造するために用いられるメチルトランスフェラーゼとしては、テアフラビンのガロイル基中の水酸基をメチル化できる酵素であれば特に限定されるものではなく、フェノール性水酸基をメチル化することが可能な一般的なメチルトランスフェラーゼの中から適宜選択して用いることができる。このようなメチルトランスフェラーゼとして、例えば、微生物由来や植物体由来のメチルトランスフェラーゼ、ラット、豚、牛肝臓由来のカテコールメチルトランスフェラーゼ等が挙げられる。
【0039】
本発明のメチル化テアフラビンを製造するためには、担子菌類の子実体又は菌糸体から得たメチルトランスフェラーゼを利用することが好ましく、エノキタケ由来メチルトランスフェラーゼ又は当該酵素の他の担子菌類由来のホモログであることがより好ましく、エノキタケ由来メチルトランスフェラーゼであることが特に好ましい。
【0040】
エノキタケ由来メチルトランスフェラーゼの他の担子菌類由来のホモログは、具体的には、配列番号2で表されるアミノ酸配列と40%以上、好ましくは50%以上、より好ましくは55%以上、さらに好ましくは62%以上の相同性を有し、かつ、メチルトランスフェラーゼ酵素活性を有するポリペプチドが挙げられる。
【0041】
なお、塩基配列とアミノ酸配列の相同性検索をする場合には、一般的に、最初にGAPを考慮せずホモロジーの核となる部分を探し、そこからアライメント処理を行う方法(Homology Search)と、置き換えや欠損を考慮しながら、塩基(アミノ酸)配列間で一致する塩基(アミノ酸残基)対が最大になるように並べ替える方法(Maximum Matching)とがある。本発明及び本願明細書における「相同性」の値は、Maximum Matchingを用いて解析し、得られる値を示す。Maximum Matchingによる相同性検索は、GENETYX Version6.1.0(GENETYX社製)等の汎用されている解析ソフトウェアを用いて行うことができる。
【0042】
メチル化テアフラビンの製造に用いられるメチルトランスフェラーゼは、天然の酵素であってもよく、公知の遺伝子組換え技術等を用いて製造された組換え酵素であってもよい。組換え酵素としては、天然の酵素と同一のアミノ酸配列を有するものであってもよく、天然の酵素に対して1以上のアミノ酸を置換・欠失・付加したアミノ酸配列からなり、かつメチルトランスフェラーゼ酵素活性を保持している改変体であってもよい。また、これらのメチルトランスフェラーゼとしては、市販のものを用いてもよい。
【0043】
天然のメチルトランスフェラーゼは、当該酵素を含む生物体から抽出し、精製することにより、得ることができる。生物体からの精製方法は、特に限定されるものではなく、一般的に行われる公知の精製方法の中から、適宜選択して行うことができる。また、メチルトランスフェラーゼ酵素活性が発揮される状態まで精製すればよく、精製度は特に限定されず、粗精製であってもよい。
【0044】
例えば、担子菌類の子実体又は菌糸体由来メチルトランスフェラーゼの場合、具体的には、本発明の遺伝子を含む担子菌類の子実体又は菌糸体を、適当な緩衝液中で超音波処理等により破砕した後、回収された上清を粗酵素とすることができる。その他、担子菌類の子実体又は菌糸体を、適当な緩衝液中でホモジナイズした後、上清を回収してもよい。粗酵素を得るための緩衝液としては、例えば、pH6.5〜8.5のトリス緩衝液やリン酸緩衝液等を用いることができる。
【0045】
また、酵素を抽出する担子菌類は、採取直後のものであってもよく、市販等されているものであってもよく、培養物であってもよい。なお、担子菌類の菌糸体の培養は、常法により行うことができる。
【0046】
このようにして得られた粗酵素は、そのまま酵素反応に用いることができる。さらには、遠心分離、限外ろ過膜、塩析、各種クロマトグラフィーを組み合わせることにより、粗酵素から精製物を得ることもできる。
【0047】
メチルトランスフェラーゼの組換え酵素は、公知のいずれの手法で製造してもよい。具体的には、メチルトランスフェラーゼをコードする遺伝子を発現ベクターに組み込み、組換え発現ベクターを作製し、この組換え発現ベクターを、微生物や培養細胞等の適当な生物体(宿主)に導入し、当該組換え発現ベクターを含む形質転換体を製造する。このようにして得られた形質転換体をそれぞれの生物体の最適な条件で培養することにより、当該形質転換体内にメチルトランスフェラーゼの組換え酵素が生成される。そこで、形質転換体の培養物から抽出し、精製することにより、メチルトランスフェラーゼの組換え酵素を得ることができる。
【0048】
酵素反応により得られたメチル化テアフラビンは、酵素反応液から、メチル化テアフラビンを抽出することが可能な溶媒等を用いることにより抽出することができる。なお、酵素反応液は、濃縮した後に抽出に用いてもよい。その他、酵素反応液を、合成吸着剤を充填したカラムに供して、メチル化テアフラビンを当該合成吸着剤に吸着させ、さらに洗浄した後、適当な溶媒を用いて溶出させることによっても、メチル化テアフラビンを得ることが可能である。さらにHPLC(High Performance LiquidChromatography)を用いて精製することにより、純度の高い新規なメチル化テアフラビンを製造することも可能である。
【0049】
本発明のメチル化テアフラビンは、飲食品、医薬品、医薬用部外品及び化粧品を始めとする様々な製品に用いることができる。本発明のメチル化テアフラビンは、これらの製品にそのまま原料として用いてもよく、その他の成分と予め混合することにより得られた組成物を原料として用いてもよい。
【0050】
本発明のメチル化テアフラビンは、非メチル化体よりも優れたpH安定性及び代謝安定性を有している。このため、本発明のメチル化テアフラビンを各種製品に用いた場合には、非メチル化体を用いる場合と比較して、より少ない添加量で、同等の機能を発揮させることができる。
【0051】
なお、本発明及び本願明細書において、pH安定性とは、pHが中性付近の溶液中で保存した場合の分解の受け難さ(安定性)をいう。また、代謝安定性とは、生体内若しくはそれに類する環境下で保存した場合の分解の受け難さをいう。
【0052】
また、本発明のメチル化テアフラビンは、脂肪蓄積抑制作用や抗酸化作用を有する。このため、本発明のメチル化テアフラビン又はこれを含む組成物を有効成分とすることにより、脂肪蓄積抑制剤や抗酸化剤を製造することができる。なお、脂肪蓄積抑制剤や抗酸化剤に含まれるメチル化テアフラビンは、1種類であってもよく、2種類以上を組み合わせて含ませてもよい。
【0053】
脂肪蓄積抑制剤としては、特に式(I)−1で表される3,5diMeTF3Gを有効成分として含むことが好ましい。3,5diMeTF3Gは、非メチル化体であるTF3Gに比べて非常に良好な脂肪蓄積抑制作用を有するためである。脂肪蓄積抑制剤に含有させるメチル化テアフラビンの量は、脂肪蓄積抑制能が発揮される濃度であれば特に限定されるものではなく、メチル化テアフラビンの種類、所望する脂肪蓄積抑制の強度等を考慮して適宜設定することができる。例えば、3,5diMeTF3Gを用いた場合には、0.5〜10μM、好ましくは0.5〜5μM、より好ましくは1〜3μMとすることができる。
【0054】
抗酸化剤としては、特に式(II)−1で表される3MeTF3’G、式(II)−3で表される3,5diMeTF3’G、式(III)−1で表される3MeTF3,3’G、式(III)−2で表される4MeTF3,3’G、又は式(III)−3で表される3,5diMeTF3,3’Gの少なくとも1種類を有効成分として含むことが好ましい。これらのメチル化テアフラビンは、対応する非メチル化体に比べて非常に良好な抗酸化作用を有するためである。抗酸化剤に含有させるメチル化テアフラビンの量は、抗酸化能が発揮される濃度であれば特に限定されるものではなく、メチル化テアフラビンの種類、所望する抗酸化能の強度等を考慮して適宜設定することができる。例えば、3MeTF3’G、3,5diMeTF3’G、3MeTF3,3’G、4MeTF3,3’G、又は3,5diMeTF3,3’Gを用いた場合には、0.1μM以上、好ましくは0.1〜50μM、より好ましくは0.1〜20μM、さらに好ましくは0.2〜2μMとすることができる。
【0055】
飲食品、医薬品、医薬用部外品及び化粧品に用いる場合には、エノキタケ等の食用担子菌類から回収されたメチルトランスフェラーゼ酵素を用いて製造されたメチル化テアフラビンを用いることが好ましい。エノキタケ等の食用担子菌類から回収されたメチルトランスフェラーゼ酵素は安全性が高いため、飲食品や医薬品、化粧品等の十分な安全性が要求される製品の工業生産に適している。また、担子菌類は簡便に大量培養(栽培)することができる。
【0056】
当該メチル化テアフラビン又はその組成物が原料として添加される飲食品としては、清涼飲料、炭酸飲料、栄養飲料、果実飲料、乳酸飲料等飲料(これらの飲料の濃縮原液及び調整用粉末を含む);アイスクリーム、アイスシャーベット、かき氷等の冷菓;そば、うどん、はるさめ、ギョーザの皮、シュウマイの皮、中華麺、即席麺等の麺類;飴、キャンディー、ガム、チョコレート、錠果、スナック菓子、ビスケット、ゼリー、ジャム、クリーム、焼き菓子等の菓子類;かまぼこ、ちくわ、ハム、ソーセージ等の水酸・畜産加工食品;加工乳、発酵乳等の乳製品;マーガリン、マヨネーズ、ショートニング、ホイップクリーム、ドレッシング等の油脂加工食品;ソース、タレ等の調味料;スープ、シチュー、サラダ、惣菜、漬物;その他種々の形態の健康食品、栄養補助食品、特定保健用食品等を挙げることができる。
【0057】
本発明のメチル化テアフラビン又はその組成物が原料として添加される医薬品及び医薬部外品の剤型としては、錠剤、液剤、カプセル剤、ドリンク剤、トローチ等が挙げられる。
また、化粧品としては、洗顔クリーム、化粧水、パック、美容液等の基礎化粧品類;ファンデーション、口紅、アイカラー等のメイクアップ化粧品類;ネイルエナメル、石鹸、入浴剤、サンスクリーン剤、デオドラントスプレー等のボディ化粧品類;シャンプー、リンス、ヘアートリートメント、ヘアームース等の頭髪用化粧品類;育毛剤、養毛剤、ヘアトニック等の頭髪用化粧品類;香水、オーデコロン等の芳香用化粧品類を挙げることが出来る。
【0058】
これらを製造するにあたり、本発明のメチル化テアフラビン又はその組成物は、通常用いられている補助的な原料や添加物と共に添加することができる。このような原料及び添加物としては、例えば、ブドウ糖、果糖、ショ糖、マルトース、ソルビトール、ステビオサイド、ルブソサイド、コーンシロップ、乳糖、L−アスコルビン酸、dl−α−トコフェロール、エリソルビン酸ナトリウム、グリセリン、プロピレングリコール、グリセリン脂肪酸エステル、ショ糖脂肪酸エステル、ソルビタン脂肪酸エステル、アラビアガム、カラギーナン、カゼイン、ゼラチン、ペクチン、寒天、ビタミンC、ビタミンB群、ビタミンE、ニコチン酸アミド、パントテン酸カルシウム、アミノ酸類、カルシウム塩類、界面活性剤、色素、香料、保存料等が挙げられる。
【実施例】
【0059】
次に参考例及び実施例を示して本発明をさらに詳細に説明するが、本発明は以下の参考例及び実施例に限定されるものではない。
【0060】
[参考例1]
エノキタケ由来メチルトランスフェラーゼ遺伝子の単離・同定を行った。
<メチルトランスフェラーゼ酵素活性のスクリーニング>
食用として市販されている日本産のシイタケ、シメジ、マイタケ、エノキタケ、ブナシメジ、ヒラタケ、ナラタケ、タモギタケ、エリンギ、アワビタケをスクリーニングに用いた。
これらの子実体を用いて柄の部分をカットし、0.5%次亜塩素酸に浸した後、滅菌水で洗浄した。柄の内部から5mm程度を切り出し、Difco Potato Dextrose Ager培地上で、25℃にて培養し、菌糸体を単離した。得られた菌糸体を、菌糸体培養用液体培地(0.02% glucose、0.01% peptone、0.002% Yeast Extract、0.002% KH2PO4、0.001% MgSO4・7H2O)に接種し、28℃で旋回培養した。
【0061】
得られた菌糸体培養液をろ過して菌糸体を回収し、粗酵素溶解液(20mM Tris−HCl(pH7.5)、1mM DTT、1mM EDTA、10% glycerol)を加えて超音波破砕した後、遠心分離して上清を回収した。この上清を粗酵素液として酵素活性測定に用いた。
酵素活性測定は、基質としてEGCGを用い、そのメチル化体の生成量を指標とした。
酵素反応液の組成は、20mM Tris−HCl(pH7.5)、2.5mM MgCl2、0.25mM EGCG、0.5mM SAM、及び50% 粗酵素液とし、全量3mlを37℃、16時間反応させた。
反応後、酵素反応液3mlに、1N HCl 70μl及び酢酸エチル5mlを加えて攪拌し、遠心分離して有機層を回収した。有機層を窒素で乾固した後、1%(w/v)アスコルビン酸含有30%メタノール水溶液に溶解し、HPLCを用いて測定した。HPLC条件を以下に示す。
【0062】
カラム:Wakopak Navi C18−5(4.6×150mm)及びC18−5(4.6×10mm)(和光純薬社製)
移動相A:蒸留水、アセトニトリル、リン酸を400:10:1(v/v)で混合した溶液移動相B:移動相Aとメタノールを2:1(v/v)で混合した溶液
グラジエント条件:20%B液(2分間)→80%B液(25分間)→80%B液(10分間)、直線濃度勾配
流速:1ml/min
検出:UV280nm
【0063】
その結果、エノキタケ培養菌糸体から抽出した粗酵素を用いた場合に、エピガロカテキン−3−O−(3−O−メチル)ガレート、エピガロカテキン−3−O−(4−O−メチル)ガレート、エピガロカテキン−3−O−(3,4−O−ジメチル)ガレート、エピガロカテキン−3−O−(3,5−O−ジメチル)ガレート、エピ(4−O−メチル)ガロカテキン−3−O−(3,5−O−ジメチル)ガレートの生成が確認された。なお、エピガロカテキン−3−O−(3,4−O−ジメチル)ガレート、エピガロカテキン−3−O−(3,5−O−ジメチル)ガレート、エピ(4−O−メチル)ガロカテキン−3−O−(3,5−O−ジメチル)ガレートに関しては、それらのピークを分取し、TOF−MS、NMRを用いて構造を確認した。また、粗酵素液を煮沸した後に酵素反応液に添加した場合や、酵素反応液にEGCGやSAMを添加しなかった場合には、これらのメチル化体の生成は認められなかった。
以上の結果から、エノキタケ培養菌糸体から抽出した粗酵素がメチルトランスフェラーゼ酵素活性を有することが確認された。
【0064】
<各種エノキタケ酵素活性の確認>
エノキタケの採取地の違い等によるメチルトランスフェラーゼ酵素活性の有無を確認した。エノキタケの菌糸は、農業生物資源ジーンバンクより購入したものを使用し、Difco Potato Dextrose Ager培地上で25℃にて培養した。菌糸体の液体培養及び酵素活性測定は、上記<メチルトランスフェラーゼ酵素活性のスクリーニング>と同様にして行った。試験には、MAFF番号430204、430205、430206、430207、430209、435210、430212、430214、430224、435085、430211、430213、435121、440110、440111、及び440118を用いた。その結果、それぞれの菌糸によって酵素活性の強弱はあるものの、全ての菌糸において、メチルトランスフェラーゼ酵素活性を確認した。
【0065】
<エノキタケ由来メチルトランスフェラーゼの同定>
上記と同様にしてエノキタケ菌糸体を液体培養し、得られた菌糸体培養液をろ過して菌糸体を回収し、凍結乾燥した。凍結乾燥物約12gを乳鉢で破砕した後、粗酵素溶解液600mlに懸濁した。この懸濁物を超音波破砕した後、遠心分離(10,000rpm×10min、4℃)し、回収した上清を再度遠心分離(30,000rpm×30min、4℃)して上清を回収した。この上清に、60%飽和になるように硫酸アンモニウムを添加し、攪拌、遠心分離して上清を回収した。さらに、この回収した上清に、80%飽和になるように硫酸アンモニウムを添加し、攪拌、遠心分離して沈殿物を得た。得られた沈殿物を、粗酵素溶解液40mlに溶解した後、PD−10(GEヘルスケアバイオサイエンス社製)を用いて脱塩した。脱塩したサンプルは、トリス緩衝液(20mM Tris−HCl(pH7.5)、1mM DTT)で平衡化した陰イオン交換カラム(HiPrep 16/10 DEAE FF、GEヘルスケアバイオサイエンス社製)に吸着させ、上記トリス緩衝液で調整した0〜500mM NaCl溶液の直線濃度勾配を用いて溶出し、メチルトランスフェラーゼ酵素活性が確認されたフラクションを、活性画分として分取した。なお、メチルトランスフェラーゼ酵素活性の確認は、<メチルトランスフェラーゼ酵素活性のスクリーニング>と同様にして行った。
【0066】
得られた活性画分を、限外ろ過カラムで脱塩、濃縮した後、上記トリス緩衝液で平衡化した陰イオン交換カラム(HiLoad 26/10 Q−sepharose HP、GEヘルスケアバイオサイエンス社製)に吸着させ、上記トリス緩衝液で調整した0〜500mM NaCl溶液の直線濃度勾配を用いて溶出し、活性画分を分取した。得られた活性画分を、限外ろ過カラムにて脱塩、濃縮し、再度同じ陰イオン交換カラムに吸着させ、分取画分を前条件より詳細にして活性画分を分取し、同様に限外ろ過カラムにて脱塩、濃縮を行った。
【0067】
得られた濃縮画分を、150mM NaClを含むトリス緩衝液(20mM Tris−HCl(pH7.5)、1mM DTT、150mM NaCl)で平衡化したゲルろ過カラム(HiLoad 16/60 Superdex 200 prep grade、GEヘルスケアバイオサイエンス社製)を用いて分画した。分取した各画分をSDS−PAGE電気泳動にかけ、得られたタンパク質染色像と、各画分のメチルトランスフェラーゼ酵素活性とを比較したところ、酵素活性と相関したバンドが、約24〜25kDaの位置に確認された。酵素活性が確認された画分を、さらに、上記トリス緩衝液で平衡化した陰イオン交換カラム(TSK−GEL BIOASSIST Q、東ソー社製)に吸着させ、上記トリス緩衝液で調整した0〜500mM NaCl溶液の直線濃度勾配を用いて溶出し、活性画分を分取した。活性画分を、再度SDS−PAGE電気泳動にかけた結果、約24〜25kDaの位置に再度特異的なバンドを確認した。この酵素タンパク質の内部配列を明らかにするために上記陰イオン交換カラム(HiLoad 26/10 Q−sepharose HP、GEヘルスケアバイオサイエンス社製)で精製したサンプルをSDS−PAGE電気泳動にかけ、バンドをゲルから回収した。
【0068】
回収したバンドは、常法に従ってゲル内トリプシン消化処理を行い、LC/MS/MS(サーモフィッシャーサイエンティフィック社製)を用いてタンパク質の内部配列を解析した。さらに、同じ活性画分を二次元電気泳動で電気泳動し(一次元目:pH3〜10の等電点分離、二次元目:4〜20%グラジエントのポリアクリルアミドゲル)、分子量約24〜25kDa、pH5付近のスポットをゲルから回収した。図3は、二次元電気泳動像である。図中、丸で囲んだスポットを、目的の酵素タンパク質として回収した。
【0069】
この回収したスポットに対して常法に従ってゲル内トリプシン消化処理を行い、LC/MS/MS(サーモフィッシャーサイエンティフィック社製)を用いて、スポットに存在しているタンパク質の内部配列の確認を行った。その結果、内部配列RVLEVGTLGGYSTTWLARA(配列番号3)及びTGGIIIVDNVVR(配列番号4)を得た。このアミノ酸配列をもとに、NCBIのデータベースによるホモロジー検索を行った結果、既知のO−メチルトランスフェラーゼの内部配列と高い相同性を有していた。
【0070】
<エノキタケ由来メチルトランスフェラーゼ遺伝子の単離・同定>
上記内部配列の配列情報に基づいてディジェネレートプライマーを設計し、エノキタケから回収したtotal−RNAから、エノキタケ由来メチルトランスフェラーゼ遺伝子を単離・同定した。
具体的には、まず、エノキタケ培養菌糸体約1gを、乳鉢を用いて液体窒素下で破砕したものから、TRI Reagent(シグマ社製)を用いてtotal−RNAを回収した。得られたtotal−RNA約4μgから、サーモスクリプトRT−PCRシステム(インビトロジェン社製)を用いて、55℃、50分間反応させることによりcDNAを合成した。このcDNAを鋳型とし、上記内部配列の配列情報に基づいて設計されたディジェネレートプライマー(FVOMT−F及びFVOMT−R)を用いてPCRを行い、エノキタケ由来メチルトランスフェラーゼ遺伝子の単離を行った。設計されたディジェネレートプライマーの塩基配列及びPCR条件を下記に記す。なお、塩基配列中、Sはグアニン又はシトシンを、Mはアデニン又はシトシンを、Yはチミン又はシトシンを、Dはアデニン、グアニン、又はチミンを、Kはグアニン又はチミンを、Vはアデニン、グアニン、又はシトシンを、Rはグアニン又はアデニンを、それぞれ示す。
【0071】
FVOMT−F:GAGGTSGGMACYYTDGGMGGSTA
FVOMT−R:GCKSACVACRTTRTCMAC
PCR条件:94℃,5min→(94℃,1min→55℃,1min→72℃,1min)×40cycles→72℃,7min
【0072】
PCR産物をアガロースゲル電気泳動にかけた結果、約400bpの増幅バンドが確認された。アガロースゲルから当該バンドを切り出し、QIAquick Gel Extraction Kit(キアゲン社製)を用いてPCR産物を回収し、pGEM−Tベクター(プロメガ社製)へクローニングした後、大腸菌JM−109株(タカラ社製)へ形質転換した。得られた形質転換体を、LB培地にて37℃で終夜振とう培養し、得られた培養物からQIAprep Spin Miniprep Kit(キアゲン社製)を用いてプラスミドを抽出した。抽出されたプラスミドのインサートの塩基配列を、Big Dye Terminator v3.1 Cycle Sequencing Kit(アプライドバイオシステムズ社製)及びABI PRISM 3100−AVANT Genetic Analyzer(アプライドバイオシステムズ社製)により確認した。
【0073】
確認された遺伝子の部分塩基配列をもとに、特異的プライマー(FVOMT−5’GSP1、FVOMT−5’GSP2、FVOMT−3’GSP1、FVOMT−3’GSP2)を設計し、5’側と3’側の全長を取得するためにRACE−PCR法を行った。設計した特異的プライマーの塩基配列を下記に示す。
【0074】
FVOMT−5’GSP1:AGCCTCTTAGCCTCAACAAAGTA
FVOMT−5’GSP2:TCTTCGAGCTCGAAGGTGAT
FVOMT−3’GSP1:ATCACCTTCGAGCTCGAAGA
FVOMT−3’GSP2:TACTTTGTTGAGGCTAAGAGGCT
【0075】
5’RACE−PCRは、まず、単離したtotal−RNA2.5μgから、FVOMT−5’GSP1プライマーを用い、サーモスクリプトRT−PCRシステム(インビトロジェン社製)を用いて、55℃、50分間反応させてcDNAを合成した。次いで、得られたcDNAから、5’RACE System for Rapid Amplification of cDNA Ends,Version 2.0(インビトロジェン社製)を用い、FVOMT−5’GSP1プライマー及びFVOMT−5’GSP2プライマーを用いて5’側の単離を行い、塩基配列を確認した。
同様に、3’RACE−PCRは、サーモスクリプトRT−PCRシステム(インビトロジェン社製)を用いてcDNAを合成し、3’RACE System for Rapid Amplification of cDNA Ends,Version 2.0(インビトロジェン社製)を用いて、得られたcDNAからFVOMT−3’GSP1プライマー及びFVOMT−3’GSP2プライマーを用いて3’側の単離を行い、塩基配列を確認した。
【0076】
確認された5’末端及び3’末端の塩基配列に基づき、下記に示すFVOMT−5’NdeIプライマー及びFVOMT−3’BamHIプライマーを設計し、RT−PCRを行った。得られたPCR産物をアガロースゲル電気泳動し、増幅バンドを回収した。回収したPCR産物をpGEM−Tベクターへクローニングし、大腸菌JM109株へ形質転換後、インサートの塩基配列を確認した。その結果、エノキタケ由来メチルトランスフェラーゼ酵素の全遺伝子を単離するにいたった。同定されたエノキタケ由来メチルトランスフェラーゼ遺伝子の塩基配列は、配列番号1で表される塩基配列であり、配列番号2で表されるアミノ酸配列からなるポリペプチドをコードしている。
【0077】
FVOMT−5’NdeI:TACATATGTCCAACCCGACAAGCATACT
FVOMT−3’BamHI:TAGGATCCAAGTTTGATAGCGTACAAGAATCC
【0078】
[参考例2]
<組換えエノキタケ由来メチルトランスフェラーゼ酵素の製造>
参考例1で単離されたエノキタケ由来メチルトランスフェラーゼ遺伝子を組み込んだ組換え発現ベクターを作製し、この組換え発現ベクターを大腸菌に導入して、大腸菌内で発現させたエノキタケ由来メチルトランスフェラーゼ酵素を回収した。
具体的には、まず、参考例1で作製したエノキタケ由来メチルトランスフェラーゼ遺伝子含有pGEM−Tベクターから、制限酵素NdeIとBamHIで切断し、アガロース電気泳動により、インサートを回収した。このインサートを、pET28a(+)ベクター(ノバジェン社製)のNdeI、BamHIサイトへクローニングすることにより、組換え発現ベクターを作製した。この組換え発現ベクターを大腸菌BL21(DE3)株(ストラタジーン社製)へ導入することにより、当該組換え発現ベクターを含む形質転換体を得た。
【0079】
得られた形質転換体をLB培地にて37℃で終夜振とう培養した後、その一部を再度新しいLB培地へ添加して培養した。O.D.600=0.6付近となるように培養した後、IPTGを最終濃度1mMになるように添加し、さらに28℃で振とう培養して、ヒスチジンタグ付酵素タンパク質の発現誘導を行った。発現誘導した大腸菌を遠心分離し、沈殿物を上記トリス緩衝液に懸濁した。この大腸菌懸濁液を超音波破砕し、再度遠心分離した。得られた上清(ライセート)を、12%SDS−PAGEにて電気泳動した結果、IPTG誘導前の大腸菌を同様に処理して電気泳動した場合と比較して、約29kDa付近に、IPTG処理により発現が誘導されたタンパク質のバンドを確認した。
【0080】
図4は、IPTG処理前の大腸菌のライセート(レーン1)、IPTG処理後の大腸菌のライセート(レーン2)、IPTG処理後の大腸菌のライセートからヒスチジンタグを用いて得られた精製物(レーン3)のSDS−PAGE電気泳動像である。図中、「M」は分子量マーカーを流したレーンである。図中の矢印の位置のバンド(約29kDa)を画像解析し、タンパク質量を数値化して比較した。画像解析には、Photoshop(アドビシステムズ社製)及びScion Image(Scion Corporation社製)ソフトウェアを使用した。その結果、この約29kDaのタンパク質量は、IPTG処理前を0%とした場合、IPTG処理後で8.4%、処理後の精製物で25.1%であり、当該タンパク質が、IPTG処理により発現が誘導されたヒスチジンタグを有するタンパク質であることが確認された。
【0081】
pET28a(+)ベクターのNdeI、BamHIサイトへクローニングした場合、約5kDaのベクター由来発現タンパク質(ヒスチジンタグを含む)が付加される。また、配列番号2のアミノ酸配列から算出された本酵素タンパク質の理論分子量は、24.7kDaである。つまり、図4の矢印で示した約29kDaのタンパク質は、組換え発現ベクターにより導入したアミノ酸配列から得られる推定分子量と一致し、ヒスチジンタグ付きエノキタケ由来メチルトランスフェラーゼ酵素であることが分かった。
【0082】
[参考例3]
<組換えエノキタケ由来メチルトランスフェラーゼ酵素の酵素活性>
参考例2で得られたIPTG処理後の大腸菌のライセートを、エノキタケ由来メチルトランスフェラーゼ酵素の組換え酵素の粗酵素液とし、この粗酵素液のメチルトランスフェラーゼ酵素活性を調べた。
具体的には、20mM Tris−HCl(pH7.5)、2.5mM MgCl2、0.25mM EGCG、0.5mM SAM、及び50% 粗酵素液となるように、酵素反応液3mlを調製し、この酵素反応液を37℃、16時間インキュベートし、反応させた。反応後の酵素反応液を、参考例1の<メチルトランスフェラーゼ酵素活性のスクリーニング>と同様に処理して分析した結果、エピガロカテキン−3−O−(3−O−メチル)ガレート、エピガロカテキン−3−O−(4−O−メチル)ガレート、エピガロカテキン−3−O−(3,4−O−ジメチル)ガレート、エピガロカテキン−3−O−(3,5−O−ジメチル)ガレート、エピ(4−O−メチル)ガロカテキン−3−O−(3,5−O−ジメチル)ガレートの生成が確認できた。
これらの結果から、参考例2において大腸菌内で発現させた組換え酵素が、メチルトランスフェラーゼ酵素活性を有していることが確認された。
【0083】
[参考例4]
<菌糸体抽出粗酵素液を用いたメチル化体の製造>
参考例1と同様にして、エノキタケ菌糸体培養液の菌糸体から抽出し、得られた抽出物の60%〜80%硫酸アンモニウム画分を脱塩処理したものを、エノキタケ由来メチルトランスフェラーゼ酵素の天然型酵素の粗酵素液とした。
具体的には、まず、エノキタケ菌糸体培養液をろ過して菌糸体を回収し、凍結乾燥した。得られた凍結乾燥物を乳鉢にて破砕した後、上記粗酵素溶解液に懸濁して、超音波破砕し、遠心分離して上清を回収した。この上清に、60%飽和になるように硫酸アンモニウムを添加し、攪拌、遠心分離して上清を回収した。さらに、この回収した上清に、80%飽和になるように硫酸アンモニウムを添加し、攪拌、遠心分離して沈殿物を得た。得られた沈殿物を、粗酵素溶解液に溶解した後、PD−10(GEヘルスケアバイオサイエンス社製)を用いて脱塩した。この脱塩サンプルを、エノキタケ由来メチルトランスフェラーゼ酵素の天然型酵素の粗酵素液とした。
得られた粗酵素液の各種基質に対するメチルトランスフェラーゼ活性を確認した。酵素反応液の組成は、20mM Tris−HCl(pH7.5)、2.5mM MgCl2、0.05mM 基質、0.5mM SAM、0.04%(w/v) アスコルビン酸、及び粗酵素液(全量3mlに対して0.25ml)とし、全量3mlを37℃、6時間反応させた。基質には、エピカテキン−3−O−ガレート(ECG)、カテキン−3−O−ガレート(CG)、ガロカテキン−3−O−ガレート(GCG)、カフェ酸、クロロゲン酸、エラグ酸、ブテイン、スルフレチン、ルテオリン、ミリセチン、又はロスマリン酸を、それぞれ用いた。
反応後の酵素反応液を、参考例1<メチルトランスフェラーゼ酵素活性のスクリーニング>と同様に処理して、当該酵素反応液中のメチル化体を、下記の条件でLC−MSを用いて分析した。標品と比較分析が可能なものについては同定も行った。HPLC条件及びMS条件を以下に示す。
【0084】
・HPLC条件
カラム:Inertsil ODS−3、2.1×150mm(GLサイエンス社製)
移動相A:0.1(v/v)%ギ酸水溶液
移動相B:0.1(v/v)%ギ酸を含むアセトニトリル
グラジエント条件(i):8%B液(20分間)→25%B液(88分間)
グラジエント条件(ii):8%B液(10分間)→50%B液(31分間)
流速:0.2ml/min
検出:UV280nm
【0085】
・MS条件
検出器:API3000(アプライドバイオシステムズ社製)
イオン源:ESI(ネガティブ)
カーテンガス:10
ネブライザーガス:14
ターボガス:6l/min
イオンスプレー電圧:−4000V
イオンスプレー温度:500度
コーン電圧:−41V
【0086】
LC−MS分析の結果を表1に示す。この結果、用いた全ての基質に対して、メチル化体の生成が確認された。これらの結果から、本発明の酵素は、水酸基を有する各種化合物を基質とし、メチル基を修飾することが可能であることが明らかとなった。
また、ECGのように、化合物中に複数の水酸基を有する化合物の中には、複数種類のメチル化体の生成が確認され、かつ、ジメチル化体も確認された。この結果から、本発明の酵素は、基質の化合物中の複数の水酸基のそれぞれを独立にメチル化可能であることも確認された。
【0087】
【表1】
【0088】
[参考例5]
<反応時間の検討>
参考例4で用いたエノキタケ由来メチルトランスフェラーゼ酵素の天然型酵素の粗酵素液(以下、天然型酵素)と、参考例2で用いたエノキタケ由来メチルトランスフェラーゼ酵素のヒスチジンタグ精製した組換え酵素液(以下、組換え酵素)とを用いて、酵素反応の反応時間を検討した。
酵素反応液の組成は、20mM リン酸緩衝液(pH7.0)、2.5mM MgCl2、0.05mM EGCG、0.5mM SAM、0.04%(w/v) アスコルビン酸、及び各粗酵素液(全量30mlに対して2.5ml)とし、全量30mlを37℃で反応させた。反応開始から5分、10分、15分、20分、25分、30分、35分、40分、45分、50分、55分、60分、2時間、 4時間、6時間、8時間、24時間後に、酵素反応液を1mlずつ分取した。これらの酵素反応液を、参考例1<メチルトランスフェラーゼ酵素活性のスクリーニング>と同様に処理して、当該酵素反応液中のメチル化体を、下記のHPLC条件で分析した。
【0089】
カラム:Inertsil ODS−3、2.1×150mm(GLサイエンス社製)
移動相A:0.1(v/v)%ギ酸水溶液
移動相B:0.1(v/v)%ギ酸を含むアセトニトリル
グラジエント条件:10%B液(10分間)→16%B液(50分間)
流速:0.2ml/min
検出:UV280nm
【0090】
各反応時間後に分取した酵素反応液中のEGCGメチル化体濃度を図5に示す。図5(A)が天然型酵素の結果であり、(B)が組換え酵素の結果である。この結果、天然型酵素では反応開始8時間後で、組換え酵素では20分後で、EGCGメチル化体の生成が最も高い値を示した。
【0091】
[参考例6]
<最適pHの検討>
参考例5において用いた天然型酵素及び組換え酵素の酵素反応における最適pHを調べた。
酵素反応液の組成は、20mMの各種緩衝液に、2.5mM MgCl2、0.05mM EGCG、0.5mM SAM、0.04%(w/v) アスコルビン酸、及び各粗酵素液(全量3mlに対して0.25ml)となるように添加して調製した。用いた緩衝液は、酢酸緩衝液(pH3.0〜5.5)、リン酸緩衝液(pH6.0〜7.0)、及びTris−HCl(pH7.5〜10)である。調製した酵素反応溶液の全量3mlを、37℃で反応させた。反応時間は、天然型酵素で6時間、組換え酵素で10分間とした。これらの酵素反応液を、参考例1<メチルトランスフェラーゼ酵素活性のスクリーニング>と同様に処理して、当該酵素反応液中のメチル化体を分析した。
各pHの酵素反応液中のEGCGメチル化体濃度を図6に示す。この結果、天然型酵素ではpH6.5〜8.5で、組換え酵素ではpH6以上で、EGCGメチル化体の生成が確認された。
【0092】
[参考例7]
<最適温度の検討>
参考例5で用いた天然型酵素及び組換え酵素の酵素反応における最適温度を調べた。
酵素反応液の組成は、20mM リン酸緩衝液(pH7.0)、2.5mM MgCl2、0.05mM EGCG、0.5mM SAM、0.04%(w/v) アスコルビン酸、及び各粗酵素液(全量3mlに対して0.25ml)とし、全量3mlを、4、10、20、30、37、42、50、又は60℃で、それぞれ反応させた。反応時間は、天然型酵素で6時間、組換え酵素で10分間とした。これらの酵素反応液を、参考例1<メチルトランスフェラーゼ酵素活性のスクリーニング>と同様に処理して、当該酵素反応液中のメチル化体を分析した。
各温度における酵素反応液中のEGCGメチル化体濃度を図7に示す。この結果、天然型酵素では10〜50℃において、組換え酵素では行った全ての温度において、EGCGメチル化体の合成が確認された。中でも、30〜42℃で、両酵素のいずれにおいても良好な酵素活性が確認された。
【0093】
[参考例8]
<基質となるテアフラビン標準品の調製>
島根県出雲産の紅茶(商品名:出雲国産紅茶(リーフ)、西製茶所製)を粉砕し、40倍量の2%(w/v)アスコルビン酸含有50%(v/v)EtOH水溶液を用いて、20分間の室温抽出を3回行った。抽出液は遠心分離を行い、抽出液から茶葉を取り除いた。抽出液は、減圧濃縮により有機溶媒を除去した後、Sep−Pak Vac C18(ウォーターズ社製)に供し、15%、40%、100%EtOHステップワイズ溶出を行った。テアフラビンが含まれる40%EtOH画分を凍結乾燥させ、適宜40%EtOHに再溶解し、下記の条件によりテアフラビンの分取を行った。分取したテアフラビンはSep−Pak Vac C18を用いて脱酸し、凍結乾燥品を作製した。これにより、TF3G標準品、TF3’G標準品、TF3,3’G標準品が得られた。
【0094】
分取条件;
カラム:Inertsil ODS−3(20 x 250mm、GLサイエンス社製)移動相A:0.05(v/v)%リン酸水溶液
移動相B:12.5(v/v)%酢酸エチルを含むアセトニトリル
グラジエント条件:24%B液(50分間)→50%B液(0.1分間)→50%B液(10分間)
流速:18ml/min
カラム温度:30℃
検出:UV280nm
【0095】
[実施例1]
<メチル化テアフラビンの製造>
参考例2において製造した組換え酵素を用いて、メチル化テアフラビンの製造を行った。基質としては、参考例8で得られたTF3G標準品、TF3’G標準品、TF3,3’G標準品をそれぞれ用いた。
酵素反応液の組成は、20mMリン酸緩衝液(pH6.5)、2.5mM MgCl2、0.025%(w/v)テアフラビン(基質)、0.5mM SAM、0.04%(w/v)アスコルビン酸、及びメチルトランスフェラーゼ(全量1000mlに対して33.3ml)とし、37℃、6時間で反応させた。その後、酵素反応液1000mlに対し、24mlの1N HClを加え、酵素反応を終了させた。酵素反応液に等量の酢酸エチルを加えて激しく撹拌し、静置後、酢酸エチル層を回収した。残る水層に対し、再度同様の操作を行った。回収した酢酸エチル層は減圧濃縮した後、参考例8と同じ条件でメチル化テアフラビンの分取を行った。分取したメチル化テアフラビンは、Sep−Pak C18を用いて脱酸し、凍結乾燥品を作製した。得られたメチル化テアフラビン凍結乾燥品に対して下記の条件でTOF−MS分析を行い、得られた精密分子量から分子式を算出した。また、下記の条件でNMR(H−NMR、C−NMR、HSQC、HMBC、COSY)を測定し、構造同定を行った。
【0096】
TOF−MS条件;
検出器:QSTAR ELITE(アプライドバイオシステムズ社製)
イオン源:ESI(ネガティブモード)
カーテンガス:30
イオンソースガス1:50
イオンソースガス2:50
イオンスプレー電圧:−4500V
イオンスプレー温度:450℃
デクラスタリングポテンシャル:−30V
フォーカスポテンシャル:−250
デクラスタリングポテンシャル:−15V
【0097】
NMR条件;
検出器:XWIN−NMR AV600(ブルカーバイオスピン社)
プローブ温度:300K
溶媒:アセトン−d6
【0098】
TOF−MS、及びNMR解析の結果を表2〜4に示した。既知化合物である3MeTF3Gの他に8種の新規化合物が得られた。各新規化合物をそれぞれ、3,5diMeTF3G(化学式(I)−1)、3MeTF3’G(化学式(II)−1)、4MeTF3’G(化学式(II)−2)、3,5diMeTF3’G(化学(II)−3)、3,4diMeTF3’G(式(II)−4)、3MeTF3,3’G(化学式(III)−1)、4MeTF3,3’G(化学式(III)−2)、3,5diMeTF3,3’G(化学式(III)−3)と決定した。
【0099】
【表2A】
【0100】
【表2B】
【0101】
【表3A】
【0102】
【表3B】
【0103】
【表3C】
【0104】
【表4A】
【0105】
【表4B】
【0106】
[実施例2]
<メチル化テアフラビン含有紅茶ポリフェノールの製造、及び反応効率>
標準品に代えて参考例8において調製した40%EtOH画分の凍結乾燥品を基質として、実施例1と同様に酵素反応を行った。酵素反応液は経時的に0.5mlずつ取り分け、実施例1と同様に処理した後、下記の条件で分析することにより、テアフラビンとメチル化テアフラビンの組成を調べた。
【0107】
分析条件;
カラム:Inertsil ODS−3(4.6 x 250mm、GLサイエンス社製)移動相A:0.05(v/v)%リン酸水溶液
移動相B:12.5(v/v)%酢酸エチルを含むアセトニトリル
グラジエント条件:24%B液(50分間)→50%B液(0.1分間)→50%B液(10分間)
流速:1ml/min
カラム温度:50℃
検出:UV375nm
【0108】
メチル化テアフラビン含有紅茶ポリフェノールの製造過程において、テアフラビンとメチル化テアフラビンの組成の推移を経時的に調べた結果を図8に示す。この結果、反応時間の経過とともにメチル化テアフラビンの生成量は増加し、24時間後で生成量が最大になることが明らかになった。この結果から、メチルトランスフェラーゼを用いた酵素処理によって、紅茶ポリフェノール中のメチル化テアフラビン含量を増やすことが可能であることが明らかとなった。
【0109】
[実施例3]
<メチル化テアフラビンのpH安定性評価>
実施例1で用いたテアフラビン標準品及び実施例1で得られたメチル化テアフラビンを、400μMになるように、40%(v/v)エタノール含有0.3M 酢酸緩衝液(pH5.0)又は40%(v/v)エタノール含有0.3M Tris−HCl緩衝液(pH7.5)にそれぞれ溶解し、常温におけるpH安定性を調べた。反応開始0、1、2時間後に試料を50μlずつ採取し、等量の1%(w/v)アスコルビン酸水溶液を添加して分解反応を停止させ、 40%(v/v)になるようにEtOHを加え、残存する各種テアフラビンを実施例2に記載の方法で分析した。
【0110】
反応開始2時間後のメチル化テアフラビン残存率を図9に示す。図9(A)がpH5.0の結果であり、図9(B)がpH7.5の結果である。この結果、ポリフェノールが安定とされるpH5.0では、全ての化合物がほぼ100%残存していた。一方、ポリフェノールが不安定とされるpH7.5では、メチル化テアフラビンはテアフラビンよりも良好なpH安定性を有していた。また、評価した全ての化合物において、メチル基の数が増えるほど、pH安定性が高まることが明らかになった。
【0111】
[実施例4]
<メチル化テアフラビンのヒト肝組織由来S9フラクションを用いた代謝安定性試験>
ヒト肝組織由来のS9フラクション(シグマ アルドリッチ社製)を用いて、実施例1で用いたテアフラビン標準品及び実施例1で得られたメチル化テアフラビンの代謝安定性を評価した。
反応組成は50mM Tris−HCl緩衝液(pH7.5)、0.1mM 3’−ホスホアデノシン−5’−ホスホサルフェート、1mM ウリジン−5’二リン酸−α−D−グルクロン酸、1mM ニコチンアミドアデニンジヌクレオチドリン酸、10mM MgCl2、0.063% ウシ血清アルブミン、8mM DTT、ヒト肝組織由来S9フラクション(最終量50μgタンパク質)、5μM 各種テアフラビン又はメチル化テアフラビンからなる反応液250μlを調製し、37℃でインキュベートした。反応開始0、15、30、60分後に50μlずつ分取し、45μlの氷冷したメタノールと、5μlの1%(w/v)アスコルビン酸水溶液とを加えて攪拌した。その後、14,000gで3分間遠心分離し、得られた上清から90μlを回収し、残存する各種テアフラビンを実施例2に記載の方法で分析した。
【0112】
反応開始60分後の、各種テアフラビン残存率を図10に示す。その結果、いずれのメチル化テアフラビンも、対応するテアフラビンと比較して残存率が高く、良好な代謝安定性を示した。また、評価した全ての化合物において、メチル基が増えるほどその代謝安定性が向上することが明らかになった。
【0113】
[実施例5]
<メチル化テアフラビンの脂肪蓄積抑制作用評価>
TF3Gのメチル化体の脂肪蓄積抑制作用を調べた。Tea extract(参考例8において、Sep−Pak Vac C18に供する前の抽出液)、実施例1で用いたTF3G(標準品)、実施例1で得られた3MeTF3G及び3,5diMeTF3Gを被検素材とした。
【0114】
まず、ヒト臓器脂肪由来前駆脂肪細胞(Lot.No.8F3482,三光純薬社製)を、前駆脂肪細胞用増殖培地で37℃、5%CO2インキュベーターにて培養し、増殖させた。その後、96−well plate(住友ベークライト社製)に前駆脂肪細胞を1.5×104 cells/wellとなるように撒いて24時間培養した後、培地を前駆脂肪細胞用分化培地に交換して分化を誘導した。分化誘導と同時に、最終濃度がTea extractは10μg/ml、各被検素材は1又は3μMとなるようにDMSO(ジメチルスルホキシド)に溶解して添加した。その後、1日おきに、前駆脂肪細胞用分化培地にTea extractは10μg/ml、各被検素材は1又は3μMとなるように含有させた培地に交換し、10日間培養した。培養終了後、細胞内の脂肪蓄積量をAdipoRedassay reagent(Lonza Walkersville)を用いて蛍光強度により評価した(Ex:485nm, Em:572nm)。
【0115】
各処理を施した細胞群の脂肪蓄積量の測定結果を図11に示す。図11(A)は各被検素材を1μMとなるように培地中に添加した場合の結果を、図11(B)は3μMとなるように培地中に添加した場合の結果を、それぞれ示す。この結果、TF3G、3MeTF3G、又は3,5diMeTF3Gを添加した場合には、無添加の場合やTea extractを添加した場合よりも数値が低く、脂肪蓄積が抑制されていることが明らかとなった。また、3MeTF3G及び3,5diMeTF3Gは、非メチル化体であるTF3Gよりも脂肪蓄積抑制作用が強く、中でも、3,5diMeTF3Gは、既知化合物である3MeTF3Gよりも優れた脂肪蓄積抑制作用を有することが明らかとなった。
これらの結果から、テアフラビンのうち、少なくともTF3Gにおいては、メチル化されている水酸基の数が多くなるほど、脂肪蓄積抑制作用が高まることが明らかになった。
【0116】
[実施例6]
<メチル化テアフラビンの抗酸化作用評価>
ヒト肝癌細胞株HepG2への過酸化水素水による酸化ストレスに対する、メチル化テアフラビンの抑制効果を評価した。Tea extract(参考例8において、Sep−Pak Vac C18に供する前の抽出液)、実施例1で用いたTF3’G及びTF3,3’G(標準品)、並びに実施例1で得られた3MeTF3’G、3,5diMeTF3’G、3MeTF3,3’G、4MeTF3,3’G、及び3,5diMeTF3,3’Gを被検素材とした。これらの被検素材は、予めDMSOに溶解させた。
【0117】
HepG2細胞は、10%非働化FBS(ウシ胎児血清)、100unit/ml ペニシリン、100μg/mlストレプトマイシン含有D−MEM培地で、37℃、5%CO2インキュベーターにて継代培養した。その後、96−well plate(住友ベークライト社製)にHepG2細胞を4×104 cells/wellとなるように撒き、最終濃度がTea extractは0.6、6、60μg/ml、被検素材は0.2、2、又は20μMとなるようにDMSOに溶解して添加したD−MEM培地で、37℃、5%CO2インキュベーターで23時間培養した。培地を除去した後、各ウェルに150μlの12mM 過酸化水素水含有D−MEM培地を添加し、細胞を2時間、過酸化水素水に暴露した。その後、上清を除去した各ウェルに、200μlの10%アラマーブルー含有D−MEM培地を添加し、3時間培養後、蛍光強度を測定した(Ex:544nm, Em:590nm)。
【0118】
過酸化水素水を添加しなかった場合のHepG2細胞生存率を100%とし、各濃度の被検素材を添加した場合におけるHepG2細胞生存率を求めた。HepG2細胞生存率の測定結果を図12に示す。図12(A)がTF3’G及びそのメチル化体を添加した場合の測定結果であり、図12(B)がTF3,3’G及びそのメチル化体を添加した場合の測定結果である。なお、有意差の検定はTukey法を用いた。
【0119】
過酸化水素水のみを添加した場合やTea extractを添加した場合と比較した結果、テアフラビンを添加した場合に差は確認できなかったものの、メチル化テアフラビンを添加した場合は細胞生存率がほぼ100%であり、有意に酸化ストレスが抑制されることが判明した。特に、TF3’G及びTF3,3’Gにおいては、メチル化テアフラビンのほうが、非メチル化体よりも高い抗酸化作用を有することが確認された。
なお、各被検素材を2又は20μMとなるように添加した場合には、非メチル化体と各種メチル化体の間で有意な差は得られなかった。これは、過酸化水素水濃度等の処理条件により、抗酸化作用の効果が必要十分であったためと推察される。
【0120】
なお、参考例1〜8及び実施例1〜6において使用した試薬の購入先は、以下の通りである。
Difco Potato Dextrose Ager:日本ベクトン・ディッキンソン
Peptone: 日本ベクトン・ディッキンソン
Yeast Extract:日本ベクトン・ディッキンソン
前駆脂肪細胞用増殖培地:三光純薬
D−MEM培地:和光純薬
分化培地:三光純薬
アラマーブルー:インビトロジェン
FCS:JRHバイオサイエンス
ペニシリン:GIBCO
ストレプトマイシン:GIBCO
EGCG:テアビゴ、DSMニュートリションジャパン
【0121】
エピカテキン−3−O−ガレート(ECG):フナコシ
カテキン−3−O−ガレート(CG):フナコシ
ガロカテキン−3−O−ガレート(GCG):フナコシ
カフェ酸:シグマ
クロロゲン酸:フナコシ
エラグ酸:フナコシ
ブテイン:フナコシ
スルフレチン:フナコシ
ルテオリン:フナコシ
ミリセチン:フナコシ
ロスマリン酸:フナコシ
【産業上の利用可能性】
【0122】
本発明の新規メチル化テアフラビンは、安定性及び機能性に優れているため、飲食品、医薬品、医薬部外品、化粧品等の様々な商品の製造分野において利用が可能である。
【技術分野】
【0001】
本発明は、1又は2以上の水酸基がメチル化されたガロイル基を有する新規なメチル化テアフラビンに関するものである。
【背景技術】
【0002】
植物の果実や葉等に主に含まれている成分の1つに、ポリフェノールがある。ポリフェノールとしては、例えば、フラボン類、フラボノール類、フラバノン類、イソフラボン類、アントシアニン類、フラバノール類、エラジタンニン類、フェニルプロパノイド類、アントシアニジン類、プロアントシアニジン類、カルコン類、オーロン類、フェニルエタノイド類等が挙げられる。より具体的には、没食子酸、エラグ酸、ヒドロキシチロソール、エピガロカテキン−3−O−ガレート(EGCG)、エピカテキン−3−O−ガレート(ECG)、カテキン−3−O−ガレート(CG)、ガロカテキン−3−O−ガレート(GCG)、エピカテキン(EC)、カテキン(C)、エピガロカテキン(EGC)、ガロカテキン(GC)、クロマニン、デルフィニジン、デルフィニジン3−O−グルコシド、プロシアニジンB2(PB2)、カフェ酸、クロロゲン酸、ロスマリン酸、カフェ酸フェネチルエステル、ストリクチニン、ケルセチン、イソクエルシトリン、ルチン、ミリセチン、ブテイン、スルフレチン、ルテオリン、エリオジクチオール等が挙げられる。これらのポリフェノールは、その構造に複数のフェノール性水酸基を持つことを特徴とし、抗酸化活性、血圧上昇抑制、血糖値上昇抑制、内脂肪蓄積抑制、動脈硬化抑制、抗炎症作用、抗がん性、アレルギー抑制、抗う蝕作用、消臭作用、抗菌活性等の様々な機能性が研究、報告されている。また、これらのポリフェノール以外にも、植物由来の成分に関して、様々な機能性が研究、報告されている。
【0003】
これらのポリフェノールの中でも、水酸基にメチル基が修飾されたメチル化ポリフェノールは、メチル基で修飾されていないポリフェノールに比べて、吸収性、代謝安定性、機能性等に優れているという報告が多くなされている。例えば、茶品種“青心大ぱん”、“べにほまれ”、“べにふじ”、“べにふうき”等には、エピガロカテキン−3−O−(3−O−メチル)ガレート(EGCG3”Me)やエピガロカテキン−3−O−(4−O−メチル)ガレート(EGCG4”Me)が含まれているが、これらのメチル化カテキンをマウスへ経口投与した場合の、投与60分後の血中メチル化カテキン濃度は、エピガロカテキン−3−O−ガレート(EGCG)と比較して、遊離体で約9倍高い値を示すことが報告されている(例えば、非特許文献1参照。)。また、7−ヒドロキシフラボン、7,4’−ジヒドロキシフラボン、クリシン、アピゲニンとこれらのメチル化体を比較したところ、メチル化体のほうが、Caco−2細胞を用いたヒト腸管吸収モデル試験において高い吸収性を示し、ヒト肝組織由来S9フラクションを用いた代謝安定性試験において良好な結果を示すことも報告されている(例えば、非特許文献2参照。)。さらに、マウスマスト細胞を用いたヒスタミン遊離抑制試験に関しては、EGCG3”Me、エピガロカテキン−3−O−(3,5−O−ジメチル)ガレート、及びエピ(3−O−メチル)ガロカテキン−3−O−(3,5−O−ジメチル)ガレートは、EGCGと比較して高い抗アレルギー作用を示し、かつメチル基の増加に伴ってその作用は強まる傾向を示したことも報告されている(例えば、特許文献1参照。)。
【0004】
紅茶ポリフェノールの一種であるテアフラビンにも、脂肪吸収抑制作用、抗酸化作用、血糖値上昇抑制作用や抗腫瘍作用等の様々な機能性が報告されている。これらの機能を生体内で効果的に発揮させるためには、十分量のテアフラビンを摂取することが重要だと考えられる。しかしながら、紅茶中のテアフラビン含量は少なく、かつ、酸化によって分解されやすい。このため、紅茶をそのまま飲食したとしても、テアフラビンを大量かつ安定的に摂取することは難しい。
【0005】
カテキンと同様にメチル化することにより、テアフラビンの代謝安定性や機能性を改善し得ると考えられる。実際に、紅茶においても、メチル化ポリフェノールの一種であるメチル化テアフラビンが含まれていることが報告されているが、その含量は乾燥茶葉中に約0.05%と通常のテアフラビンよりもさらに少ない(例えば、非特許文献3参照。)。
このため、メチル化テアフラビンを紅茶から大量に得ることは困難である。
【0006】
メチル化テアフラビンを人為的に得る方法としては、エピガロカテキン3−O−(3−O−メチル)ガレート等のメチル化カテキン、エピカテキン、及びエピカテキンガレートを原料として、酸化酵素により重合させる方法がある(例えば、非特許文献3参照。)。
しかしながら、天然に希少なメチル化カテキンを原料として使用するこのような方法では、そもそも原料を得ること自体が難しいことに加えて、合成されるメチル化テアフラビンは、原料となるメチル化カテキンの種類に依存する。このため、現在までに、テアフラビン3−O−(3−O−メチル)ガレート、テアフラビン3−O−(3−O−メチル)ガレート,3’−O−ガレートしか得られていない。また、非特許文献3においては、メチル化テアフラビンの機能性は明らかにされていない。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特開2008−189628号公報
【非特許文献】
【0008】
【非特許文献1】サノ(Sano)、外3名、フレグランス・ジャーナル(FRAGRANCE JOURNAL)、2000年、4号、第46〜52ページ。
【非特許文献2】ウェン(Wen)、外1名、ドラッグ・メタボリズム・アンド・ディスポジション(Drug Metabolism and Disposition)、2006年、第34巻第10号、第1786〜1792ページ。
【非特許文献3】ニシムラ(Nishimura)、外5名、ジャーナル・オブ・アグリカルチュラル・アンド・フード・ケミストリー(Journal of Agricultural and Food Chemistry) 2007年、第55巻、第7252〜7257ページ。
【発明の概要】
【発明が解決しようとする課題】
【0009】
テアフラビン等のポリフェノール類は、非常に多くの機能を有しているが、これらの機能は、ポリフェノールの種類によって異なる場合がある。同様に、メチル化ポリフェノールにおいても、ポリフェノール中のいずれの水酸基をメチル化するかによって、その作用効果や活性強度が異なる可能性が高い。このため、様々なテアフラビン類をメチル化することにより、より代謝安定性や機能性に優れたメチル化テアフラビンを得ることができると期待される。
【0010】
本発明は、上記課題を鑑みてなされたものであって、1又は2以上の水酸基がメチル化されたガロイル基を有し、安定性や機能性に優れた新規なメチル化テアフラビンを提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明者らは、上記課題を解決すべく鋭意研究した結果、テアフラビンをはじめとするポリフェノールの水酸基を効率よくメチル化することができる新規なメチルトランスフェラーゼを得、当該酵素を用いてテアフラビンを基質とした酵素反応法を行なうことにより、新規なメチル化テアフラビンを合成することにより、本発明を完成させた。
【0012】
すなわち、本発明は、
(1) 1又は2以上の水酸基がメチル化されたガロイル基を有するメチル化テアフラビンであって、下記一般式(I)〜(III)のいずれか1の式で表されることを特徴とする化合物、
【化1】
[式(I)中、R1〜R3は水素原子又はメチル基を表し、かつ、R1〜R3の少なくとも2以上がメチル基である。]
【化2】
[式(II)中、R4〜R6は水素原子又はメチル基を表し、かつ、R4〜R6の少なくとも1以上がメチル基である。]
【化3】
[式(III)中、R7〜R12は水素原子又はメチル基を表し、かつ、R7〜R12の少なくとも1以上がメチル基である。但し、R7のみがメチル基である場合を除く。]
【0013】
(2) 下記式(I)−1、(II)−1〜4、及び(III)−1〜3からなる群より選択される1の式で表されることを特徴とする、前記(1)に記載の化合物、
【化4】
【化5】
【化6】
【0014】
(3) 前記(1)又は(2)に記載の化合物を含むことを特徴とする組成物、
(4) 前記(1)又は(2)に記載の化合物を含むことを特徴とする脂肪蓄積抑制剤、(5) 前記化合物が前記式(I)−1で表される化合物であることを特徴とする前記(4)記載の脂肪蓄積抑制剤、
(6) 前記(1)又は(2)に記載の化合物を含むことを特徴とする抗酸化剤、
(7) 前記化合物が、前記式(II)−1、(II)−3、(III)−1、(III)−2、及び(III)−3で表される化合物から選択される1種以上であることを特徴とする前記(6)記載の抗酸化剤、
(8) 前記(1)又は(2)に記載の化合物を含むことを特徴とする飲食品、
(9) 前記(1)又は(2)に記載の化合物を含むことを特徴とする医薬品又は医薬部外品、
(10) 前記(1)又は(2)に記載の化合物を含むことを特徴とする化粧品、
(11) 下記式(I)−0で表される化合物、下記式(II)−0で表される化合物、及び下記式(III)−0で表される化合物からなる群より選択される1種以上の化合物を基質として、メチルトランスフェラーゼ酵素を反応させることにより、前記一般式(I)で表される化合物、前記一般式(II)で表される化合物、及び前記一般式(III)で表される化合物からなる群より選択される1種以上の化合物を合成することを特徴とする、メチル化テアフラビンの製造方法、
【化7】
(12) 前記メチルトランスフェラーゼ酵素が、エノキタケ(Flammulinavelutipes)由来の酵素であることを特徴とする前記(11)記載のメチル化テアフラビンの製造方法、
を提供するものである。
【発明の効果】
【0015】
本発明のメチル化テアフラビンは、非メチル化体であるテアフラビンと同様の生理機能を有する上に、非常に良好なpH安定性及び代謝安定性を有する。このため、本発明のメチル化テアフラビンを飲食品、医薬品、医薬部外品、化粧品等の様々な商品へ応用することにより、非メチル化体を使用するよりも、機能性や経済性に優れた製品の開発が可能となる。
【図面の簡単な説明】
【0016】
【図1】エノキタケ由来メチルトランスフェラーゼ遺伝子の塩基配列及び対応するアミノ酸配列を示した図である。
【図2】図2(A)は、配列番号2で表されるアミノ酸配列と、上記2種類の担子菌の遺伝子と、相同性検察の結果相同性が高かった上位4遺伝子とのアミノ酸配列を比較した結果である。図2(B)は、図2(A)中の1〜4に示す領域から、O−メチルトランスフェラーゼ酵素のコンセンサス配列であると推定された配列を表す図である。
【図3】参考例1において、エノキタケ菌糸培養物から分画されたメチルトランスフェラーゼ酵素活性を有する活性画分の二次元電気泳動像である。
【図4】参考例2において、IPTG処理前の大腸菌のライセート(レーン1)、IPTG処理後の大腸菌のライセート(レーン2)、IPTG処理後の大腸菌のライセートからヒスチジンタグを用いて得られた精製物(レーン3)のSDS−PAGE電気泳動像である。
【図5】参考例5において、各反応時間後に分取した酵素反応液中のEGCGメチル化体濃度を示した図である。図5(A)が天然型酵素の結果であり、図5(B)が組換え酵素の結果である。
【図6】参考例6において、各pHの酵素反応液中のEGCGメチル化体濃度を示した図である。
【図7】参考例7において、各温度の酵素反応液中のEGCGメチル化体濃度を示した図である。
【図8】実施例2において、メチル化テアフラビン含有紅茶ポリフェノールの製造過程におけるテアフラビンとメチル化テアフラビンの組成を経時的に測定した結果を示した図である。
【図9】実施例3において、各種テアフラビン及びメチル化テアフラビンの残存率を反応開始2時間後に測定した結果を示した図である。図9(A)がpH5.0の結果であり、図9(B)がpH7.5の結果である。
【図10】実施例4において、各種テアフラビン及びメチル化テアフラビンの残存率を反応開始60分後に測定した結果を示した図である。
【図11】実施例5において、各処理を施した細胞群の脂肪蓄積量の測定結果を示した図である。図11(A)は各被検素材を1μMとなるように培地中に添加した場合の結果であり、図11(B)は3μMとなるように培地中に添加した場合の結果である。
【図12】実施例6において、HepG2細胞生存率の測定結果を示した図である。図12(A)がTF3’G及びそのメチル化体を添加した場合の測定結果であり、図12(B)がTF3,3’G及びそのメチル化体を添加した場合の測定結果である。
【発明を実施するための形態】
【0017】
本発明及び本願明細書において使用する各化合物の略称を以下に示す。
TF:テアフラビン
TF3G:テアフラビン3−O−ガレート
3MeTF3G:テアフラビン3−O−(3−O−メチル)ガレート
3,5diMeTF3G:テアフラビン3−O−(3,5−O−ジメチル)ガレート
TF3’G:テアフラビン3’−O−ガレート
3MeTF3’G:テアフラビン3’−O−(3−O−メチル)ガレート
4MeTF3’G:テアフラビン3’−O−(4−O−メチル)ガレート
3,5diMeTF3’G:テアフラビン3’−O−(3,5−O−ジメチル)ガレート3,4diMeTF3’G:テアフラビン3’−O−(3,4−O−ジメチル)ガレートTF3,3’G:テアフラビン3,3’ −ジ−O−ガレート
3MeTF3,3’G:テアフラビン3−O−ガレート,3’−O−(3−O−メチル)ガレート
4MeTF3,3’G:テアフラビン3−O−ガレート,3’−O−(4−O−メチル)ガレート
3,5diMeTF3,3’G:テアフラビン3−O−ガレート,3’−O−(3,5−O−ジメチル)ガレート
Tea extract:紅茶抽出物、紅茶エキス
【0018】
本発明及び本願明細書において、テアフラビンとは、下記式(I)−0で表される化合物(TF3G)、下記式(II)−0で表される化合物(TF3’G)、又は下記式(III)−0で表される化合物(TF3,3’G)を示す。なお、これらの式で表される化合物は複数の異性体を有しているが、これらの異性体も、テアフラビンに含まれる。
また、本発明及び本願明細書において、メチル化テアフラビンとは、テアフラビンのガロイル基中の少なくとも1の水酸基がメチル化されている化合物を意味する。
【0019】
【化8】
【0020】
<新規メチルトランスフェラーゼ>
本発明者らは、新規なメチル化テアフラビンを合成するために、後記参考例1等に示すように、エノキタケ(Flammulina velutipes)由来の新規なメチルトランスフェラーゼを単離・同定した。
得られたエノキタケ由来メチルトランスフェラーゼをコードする遺伝子の塩基配列(配列番号1)及び対応するアミノ酸配列(配列番号2)を図1に示す。配列番号1で表される塩基配列の45位〜734位がコーディング領域である。
【0021】
なお、配列番号1で表される塩基配列及び配列番号2で表されるアミノ酸配列に対して、公開されているNCBI(National Center for Biotechnology Information)のデータベースを用いてホモロジー検索を行った結果、最も相同性の高い遺伝子はオオキツネタケ由来遺伝子S238N−H82(アクセッション番号:XM_001878803)であり、塩基配列で63.6%、アミノ酸配列で61.6%程度の相同性であった。
【0022】
また、全ゲノム配列が判明している担子菌類であるモデルキノコのデータベース(http://genome.jgi−psf.org/Phchr1/Phchr1.home.html)を用いて、同様にホモロジー検索を行った結果、最も相同性の高い遺伝子はO-メチルトランスフェラーゼ遺伝子(Protein ID:127115)であり、塩基配列で63.9%、アミノ酸配列で56.8%程度の相同性であった。
【0023】
図2(A)は、配列番号2で表されるアミノ酸配列と、上記2種類の担子菌の遺伝子と、相同性検察の結果相同性が高かった上位4遺伝子とのアミノ酸配列を比較した結果である。図2(A)中、「F.velutipes」は配列番号2で表されるアミノ酸配列を表す。また、「L.bicolor S238N−H82」はオオキツネタケ由来遺伝子S238N−H82、「P.chrysosporium」はモデルキノコ由来O−メチルトランスフェラーゼ遺伝子、「B.indeica subsp.」はBeijerinckia indica subsp.由来O−メチルトランスフェラーゼ ファミリータンパク質遺伝子(アクセッション番号:YP_001832797)、「B.ambifaria AMMD」はBurkholderia ambifaria AMMD由来O−メチルトランスフェラーゼ ファミリータンパク質遺伝子(アクセッション番号:YP_777310)、「B.thailandensis」はBurkholderia thailandensis MSMB43由来O−メチルトランスフェラーゼ ファミリータンパク質遺伝子(アクセッション番号:ZP_02466609)、及び「E.sakazakii」はEnterobacter sakazakii ATCC BAA−894由来遺伝子(アクセッション番号:YP_001437785)の推定アミノ酸配列をそれぞれ表す。
【0024】
本発明者らは、このアラインメントから、図2(A)中、1〜4に示す領域が、全7種類の遺伝子において保存性の高い領域であることを見出した。なお、配列番号2で表されるアミノ酸配列中、領域1は70〜88番目の領域であり、領域2は139〜148番目の領域であり、領域3は171〜175番目の領域であり、領域4は215〜223番目の領域である。さらに本発明者らは、これらの4領域のアミノ酸配列から、O−メチルトランスフェラーゼ酵素のコンセンサス配列を推定した。図2(B)は、図2(A)中の1〜4に示す領域から、O−メチルトランスフェラーゼ酵素のコンセンサス配列であると推定された配列を表す。なお、図2(B)中、「x」はいずれのアミノ酸残基であってもよいことを意味し、「(/)」は、括弧内のいずれかのアミノ酸残基であることを意味する。
【0025】
さらに、配列番号2で表されるアミノ酸配列に対して、NCBIデータベースを用いて植物由来のメチルトランスフェラーゼ酵素とのホモロジー検索を行った結果、ダイズ(Zea mays)由来遺伝子(アクセッション番号:AY279004)と、塩基配列で53.9%、アミノ酸配列で30.4%の相同性であった。同様に、イネ(Olyzasativa)由来遺伝子(アクセッション番号:AY279004)と50.7%(塩基配列)及び30.0%(アミノ酸配列)、シロイヌナズナ(Arabidopsisthaliana)由来遺伝子(アクセッション番号:MN_116065)52.3%(塩基配列)及び30.8%(アミノ酸配列)程度の相同性であった。さらに、特許文献4に記載の、茶から単離されたEGCGをメチル化可能なメチルトランスフェラーゼ酵素と比較した結果、塩基配列で53.9%、アミノ酸配列では30.4%程度の相同性であった。
【0026】
本発明者らは、テアフラビンを基質とし、同定したエノキタケ由来メチルトランスフェラーゼによる酵素反応によって、メチル化テアフラビンを得た。この結果、公知のメチル化テアフラビンである3MeTF3G以外にも、多くの新規なメチル化テアフラビンを得た。
【0027】
すなわち、本発明のメチル化テアフラビンは、下記一般式(I)〜(III)のいずれか1の式で表されることを特徴とする。
【0028】
【化9】
[式(I)中、R1〜R3は水素原子又はメチル基を表し、かつ、R1〜R3の少なくとも2以上がメチル基である。]
【0029】
【化10】
[式(II)中、R4〜R6は水素原子又はメチル基を表し、かつ、R4〜R6の少なくとも1以上がメチル基である。]
【0030】
【化11】
[式(III)中、R7〜R12は水素原子又はメチル基を表し、かつ、R7〜R12の少なくとも1以上がメチル基である。但し、R7のみがメチル基である場合を除く。]
【0031】
一般式(I)中、R1〜R3は水素原子又はメチル基を表し、かつ、R1〜R3の少なくとも2以上がメチル基である。
一般式(II)中、R4〜R6は水素原子又はメチル基を表し、かつ、R4〜R6の少なくとも1以上がメチル基である。
一般式(III)中、R7〜R12は水素原子又はメチル基を表し、かつ、R7〜R12の少なくとも1以上がメチル基である。但し、R7のみがメチル基である場合を除く。
なお、これらの一般式で表される化合物は複数の異性体を有しているが、これらの一般式で表される化合物である限り、いずれの異性体も、本発明のメチル化テアフラビンに含まれる。
【0032】
本発明のメチル化テアフラビンとしては、特に、式(I)−1で表される化合物(3,5diMeTF3G)、式(II)−1で表される化合物(3MeTF3’G)、式(II)−2で表される化合物(4MeTF3’G)、式(II)−3で表される化合物(3,5diMeTF3’G)、式(II)−4で表される化合物(3,4diMeTF3’G)、式(III)−1で表される化合物(3MeTF3,3’G)、式(III)−2で表される化合物(4MeTF3,3’G)、及び式(III)−3で表される化合物(3,5diMeTF3,3’G)であることが好ましい。
【0033】
【化12】
【0034】
【化13】
【0035】
【化14】
【0036】
本発明のメチル化テアフラビンは、エノキタケ由来メチルトランスフェラーゼを用いた酵素反応により初めて合成されたものである。紅茶にはテアフラビンが含まれており、かつ生茶葉由来のメチルトランスフェラーゼも公知である(例えば、特開2007−306806号公報)。にもかかわらず、今までこれらのメチル化テアフラビンが得られていなかった理由は明らかではないが、生茶葉由来メチルトランスフェラーゼがカテキン類に対する特異性が高く、テアフラビン類に対する特異性が十分ではない可能性や、エノキタケ由来メチルトランスフェラーゼの酵素活性が、生茶葉由来メチルトランスフェラーゼ等の公知のメチルトランスフェラーゼよりも高い可能性がある。
【0037】
本発明のメチル化テアフラビンは、テアフラビンをメチルトランスフェラーゼによりメチル化することによって製造することができる。より具体的には、例えば、1種類又は2種類以上のメチルトランスフェラーゼを、pH5〜12、好ましくはpH6〜9、より好ましくはpH6.5〜8.5の緩衝液に懸濁し、さらにテアフラビン含有物とメチル基の供与体とを加えて、0〜80℃、好ましくは10〜50℃、より好ましくは30〜42℃で反応させることにより、酵素反応液中にメチル化体を得ることができる。なお、テアフラビン含有物としては、紅茶、紅茶抽出物、紅茶ポリフェノール、テアフラビン標準品等を用いることができる。また、メチル基の供与体としては、S−adenosyl−L−methionine (SAM)等の、一般的にフェノール性水酸基のメチル化の際に用いられる物質の中から、適宜選択して用いることができる。
【0038】
本発明のメチル化テアフラビンを製造するために用いられるメチルトランスフェラーゼとしては、テアフラビンのガロイル基中の水酸基をメチル化できる酵素であれば特に限定されるものではなく、フェノール性水酸基をメチル化することが可能な一般的なメチルトランスフェラーゼの中から適宜選択して用いることができる。このようなメチルトランスフェラーゼとして、例えば、微生物由来や植物体由来のメチルトランスフェラーゼ、ラット、豚、牛肝臓由来のカテコールメチルトランスフェラーゼ等が挙げられる。
【0039】
本発明のメチル化テアフラビンを製造するためには、担子菌類の子実体又は菌糸体から得たメチルトランスフェラーゼを利用することが好ましく、エノキタケ由来メチルトランスフェラーゼ又は当該酵素の他の担子菌類由来のホモログであることがより好ましく、エノキタケ由来メチルトランスフェラーゼであることが特に好ましい。
【0040】
エノキタケ由来メチルトランスフェラーゼの他の担子菌類由来のホモログは、具体的には、配列番号2で表されるアミノ酸配列と40%以上、好ましくは50%以上、より好ましくは55%以上、さらに好ましくは62%以上の相同性を有し、かつ、メチルトランスフェラーゼ酵素活性を有するポリペプチドが挙げられる。
【0041】
なお、塩基配列とアミノ酸配列の相同性検索をする場合には、一般的に、最初にGAPを考慮せずホモロジーの核となる部分を探し、そこからアライメント処理を行う方法(Homology Search)と、置き換えや欠損を考慮しながら、塩基(アミノ酸)配列間で一致する塩基(アミノ酸残基)対が最大になるように並べ替える方法(Maximum Matching)とがある。本発明及び本願明細書における「相同性」の値は、Maximum Matchingを用いて解析し、得られる値を示す。Maximum Matchingによる相同性検索は、GENETYX Version6.1.0(GENETYX社製)等の汎用されている解析ソフトウェアを用いて行うことができる。
【0042】
メチル化テアフラビンの製造に用いられるメチルトランスフェラーゼは、天然の酵素であってもよく、公知の遺伝子組換え技術等を用いて製造された組換え酵素であってもよい。組換え酵素としては、天然の酵素と同一のアミノ酸配列を有するものであってもよく、天然の酵素に対して1以上のアミノ酸を置換・欠失・付加したアミノ酸配列からなり、かつメチルトランスフェラーゼ酵素活性を保持している改変体であってもよい。また、これらのメチルトランスフェラーゼとしては、市販のものを用いてもよい。
【0043】
天然のメチルトランスフェラーゼは、当該酵素を含む生物体から抽出し、精製することにより、得ることができる。生物体からの精製方法は、特に限定されるものではなく、一般的に行われる公知の精製方法の中から、適宜選択して行うことができる。また、メチルトランスフェラーゼ酵素活性が発揮される状態まで精製すればよく、精製度は特に限定されず、粗精製であってもよい。
【0044】
例えば、担子菌類の子実体又は菌糸体由来メチルトランスフェラーゼの場合、具体的には、本発明の遺伝子を含む担子菌類の子実体又は菌糸体を、適当な緩衝液中で超音波処理等により破砕した後、回収された上清を粗酵素とすることができる。その他、担子菌類の子実体又は菌糸体を、適当な緩衝液中でホモジナイズした後、上清を回収してもよい。粗酵素を得るための緩衝液としては、例えば、pH6.5〜8.5のトリス緩衝液やリン酸緩衝液等を用いることができる。
【0045】
また、酵素を抽出する担子菌類は、採取直後のものであってもよく、市販等されているものであってもよく、培養物であってもよい。なお、担子菌類の菌糸体の培養は、常法により行うことができる。
【0046】
このようにして得られた粗酵素は、そのまま酵素反応に用いることができる。さらには、遠心分離、限外ろ過膜、塩析、各種クロマトグラフィーを組み合わせることにより、粗酵素から精製物を得ることもできる。
【0047】
メチルトランスフェラーゼの組換え酵素は、公知のいずれの手法で製造してもよい。具体的には、メチルトランスフェラーゼをコードする遺伝子を発現ベクターに組み込み、組換え発現ベクターを作製し、この組換え発現ベクターを、微生物や培養細胞等の適当な生物体(宿主)に導入し、当該組換え発現ベクターを含む形質転換体を製造する。このようにして得られた形質転換体をそれぞれの生物体の最適な条件で培養することにより、当該形質転換体内にメチルトランスフェラーゼの組換え酵素が生成される。そこで、形質転換体の培養物から抽出し、精製することにより、メチルトランスフェラーゼの組換え酵素を得ることができる。
【0048】
酵素反応により得られたメチル化テアフラビンは、酵素反応液から、メチル化テアフラビンを抽出することが可能な溶媒等を用いることにより抽出することができる。なお、酵素反応液は、濃縮した後に抽出に用いてもよい。その他、酵素反応液を、合成吸着剤を充填したカラムに供して、メチル化テアフラビンを当該合成吸着剤に吸着させ、さらに洗浄した後、適当な溶媒を用いて溶出させることによっても、メチル化テアフラビンを得ることが可能である。さらにHPLC(High Performance LiquidChromatography)を用いて精製することにより、純度の高い新規なメチル化テアフラビンを製造することも可能である。
【0049】
本発明のメチル化テアフラビンは、飲食品、医薬品、医薬用部外品及び化粧品を始めとする様々な製品に用いることができる。本発明のメチル化テアフラビンは、これらの製品にそのまま原料として用いてもよく、その他の成分と予め混合することにより得られた組成物を原料として用いてもよい。
【0050】
本発明のメチル化テアフラビンは、非メチル化体よりも優れたpH安定性及び代謝安定性を有している。このため、本発明のメチル化テアフラビンを各種製品に用いた場合には、非メチル化体を用いる場合と比較して、より少ない添加量で、同等の機能を発揮させることができる。
【0051】
なお、本発明及び本願明細書において、pH安定性とは、pHが中性付近の溶液中で保存した場合の分解の受け難さ(安定性)をいう。また、代謝安定性とは、生体内若しくはそれに類する環境下で保存した場合の分解の受け難さをいう。
【0052】
また、本発明のメチル化テアフラビンは、脂肪蓄積抑制作用や抗酸化作用を有する。このため、本発明のメチル化テアフラビン又はこれを含む組成物を有効成分とすることにより、脂肪蓄積抑制剤や抗酸化剤を製造することができる。なお、脂肪蓄積抑制剤や抗酸化剤に含まれるメチル化テアフラビンは、1種類であってもよく、2種類以上を組み合わせて含ませてもよい。
【0053】
脂肪蓄積抑制剤としては、特に式(I)−1で表される3,5diMeTF3Gを有効成分として含むことが好ましい。3,5diMeTF3Gは、非メチル化体であるTF3Gに比べて非常に良好な脂肪蓄積抑制作用を有するためである。脂肪蓄積抑制剤に含有させるメチル化テアフラビンの量は、脂肪蓄積抑制能が発揮される濃度であれば特に限定されるものではなく、メチル化テアフラビンの種類、所望する脂肪蓄積抑制の強度等を考慮して適宜設定することができる。例えば、3,5diMeTF3Gを用いた場合には、0.5〜10μM、好ましくは0.5〜5μM、より好ましくは1〜3μMとすることができる。
【0054】
抗酸化剤としては、特に式(II)−1で表される3MeTF3’G、式(II)−3で表される3,5diMeTF3’G、式(III)−1で表される3MeTF3,3’G、式(III)−2で表される4MeTF3,3’G、又は式(III)−3で表される3,5diMeTF3,3’Gの少なくとも1種類を有効成分として含むことが好ましい。これらのメチル化テアフラビンは、対応する非メチル化体に比べて非常に良好な抗酸化作用を有するためである。抗酸化剤に含有させるメチル化テアフラビンの量は、抗酸化能が発揮される濃度であれば特に限定されるものではなく、メチル化テアフラビンの種類、所望する抗酸化能の強度等を考慮して適宜設定することができる。例えば、3MeTF3’G、3,5diMeTF3’G、3MeTF3,3’G、4MeTF3,3’G、又は3,5diMeTF3,3’Gを用いた場合には、0.1μM以上、好ましくは0.1〜50μM、より好ましくは0.1〜20μM、さらに好ましくは0.2〜2μMとすることができる。
【0055】
飲食品、医薬品、医薬用部外品及び化粧品に用いる場合には、エノキタケ等の食用担子菌類から回収されたメチルトランスフェラーゼ酵素を用いて製造されたメチル化テアフラビンを用いることが好ましい。エノキタケ等の食用担子菌類から回収されたメチルトランスフェラーゼ酵素は安全性が高いため、飲食品や医薬品、化粧品等の十分な安全性が要求される製品の工業生産に適している。また、担子菌類は簡便に大量培養(栽培)することができる。
【0056】
当該メチル化テアフラビン又はその組成物が原料として添加される飲食品としては、清涼飲料、炭酸飲料、栄養飲料、果実飲料、乳酸飲料等飲料(これらの飲料の濃縮原液及び調整用粉末を含む);アイスクリーム、アイスシャーベット、かき氷等の冷菓;そば、うどん、はるさめ、ギョーザの皮、シュウマイの皮、中華麺、即席麺等の麺類;飴、キャンディー、ガム、チョコレート、錠果、スナック菓子、ビスケット、ゼリー、ジャム、クリーム、焼き菓子等の菓子類;かまぼこ、ちくわ、ハム、ソーセージ等の水酸・畜産加工食品;加工乳、発酵乳等の乳製品;マーガリン、マヨネーズ、ショートニング、ホイップクリーム、ドレッシング等の油脂加工食品;ソース、タレ等の調味料;スープ、シチュー、サラダ、惣菜、漬物;その他種々の形態の健康食品、栄養補助食品、特定保健用食品等を挙げることができる。
【0057】
本発明のメチル化テアフラビン又はその組成物が原料として添加される医薬品及び医薬部外品の剤型としては、錠剤、液剤、カプセル剤、ドリンク剤、トローチ等が挙げられる。
また、化粧品としては、洗顔クリーム、化粧水、パック、美容液等の基礎化粧品類;ファンデーション、口紅、アイカラー等のメイクアップ化粧品類;ネイルエナメル、石鹸、入浴剤、サンスクリーン剤、デオドラントスプレー等のボディ化粧品類;シャンプー、リンス、ヘアートリートメント、ヘアームース等の頭髪用化粧品類;育毛剤、養毛剤、ヘアトニック等の頭髪用化粧品類;香水、オーデコロン等の芳香用化粧品類を挙げることが出来る。
【0058】
これらを製造するにあたり、本発明のメチル化テアフラビン又はその組成物は、通常用いられている補助的な原料や添加物と共に添加することができる。このような原料及び添加物としては、例えば、ブドウ糖、果糖、ショ糖、マルトース、ソルビトール、ステビオサイド、ルブソサイド、コーンシロップ、乳糖、L−アスコルビン酸、dl−α−トコフェロール、エリソルビン酸ナトリウム、グリセリン、プロピレングリコール、グリセリン脂肪酸エステル、ショ糖脂肪酸エステル、ソルビタン脂肪酸エステル、アラビアガム、カラギーナン、カゼイン、ゼラチン、ペクチン、寒天、ビタミンC、ビタミンB群、ビタミンE、ニコチン酸アミド、パントテン酸カルシウム、アミノ酸類、カルシウム塩類、界面活性剤、色素、香料、保存料等が挙げられる。
【実施例】
【0059】
次に参考例及び実施例を示して本発明をさらに詳細に説明するが、本発明は以下の参考例及び実施例に限定されるものではない。
【0060】
[参考例1]
エノキタケ由来メチルトランスフェラーゼ遺伝子の単離・同定を行った。
<メチルトランスフェラーゼ酵素活性のスクリーニング>
食用として市販されている日本産のシイタケ、シメジ、マイタケ、エノキタケ、ブナシメジ、ヒラタケ、ナラタケ、タモギタケ、エリンギ、アワビタケをスクリーニングに用いた。
これらの子実体を用いて柄の部分をカットし、0.5%次亜塩素酸に浸した後、滅菌水で洗浄した。柄の内部から5mm程度を切り出し、Difco Potato Dextrose Ager培地上で、25℃にて培養し、菌糸体を単離した。得られた菌糸体を、菌糸体培養用液体培地(0.02% glucose、0.01% peptone、0.002% Yeast Extract、0.002% KH2PO4、0.001% MgSO4・7H2O)に接種し、28℃で旋回培養した。
【0061】
得られた菌糸体培養液をろ過して菌糸体を回収し、粗酵素溶解液(20mM Tris−HCl(pH7.5)、1mM DTT、1mM EDTA、10% glycerol)を加えて超音波破砕した後、遠心分離して上清を回収した。この上清を粗酵素液として酵素活性測定に用いた。
酵素活性測定は、基質としてEGCGを用い、そのメチル化体の生成量を指標とした。
酵素反応液の組成は、20mM Tris−HCl(pH7.5)、2.5mM MgCl2、0.25mM EGCG、0.5mM SAM、及び50% 粗酵素液とし、全量3mlを37℃、16時間反応させた。
反応後、酵素反応液3mlに、1N HCl 70μl及び酢酸エチル5mlを加えて攪拌し、遠心分離して有機層を回収した。有機層を窒素で乾固した後、1%(w/v)アスコルビン酸含有30%メタノール水溶液に溶解し、HPLCを用いて測定した。HPLC条件を以下に示す。
【0062】
カラム:Wakopak Navi C18−5(4.6×150mm)及びC18−5(4.6×10mm)(和光純薬社製)
移動相A:蒸留水、アセトニトリル、リン酸を400:10:1(v/v)で混合した溶液移動相B:移動相Aとメタノールを2:1(v/v)で混合した溶液
グラジエント条件:20%B液(2分間)→80%B液(25分間)→80%B液(10分間)、直線濃度勾配
流速:1ml/min
検出:UV280nm
【0063】
その結果、エノキタケ培養菌糸体から抽出した粗酵素を用いた場合に、エピガロカテキン−3−O−(3−O−メチル)ガレート、エピガロカテキン−3−O−(4−O−メチル)ガレート、エピガロカテキン−3−O−(3,4−O−ジメチル)ガレート、エピガロカテキン−3−O−(3,5−O−ジメチル)ガレート、エピ(4−O−メチル)ガロカテキン−3−O−(3,5−O−ジメチル)ガレートの生成が確認された。なお、エピガロカテキン−3−O−(3,4−O−ジメチル)ガレート、エピガロカテキン−3−O−(3,5−O−ジメチル)ガレート、エピ(4−O−メチル)ガロカテキン−3−O−(3,5−O−ジメチル)ガレートに関しては、それらのピークを分取し、TOF−MS、NMRを用いて構造を確認した。また、粗酵素液を煮沸した後に酵素反応液に添加した場合や、酵素反応液にEGCGやSAMを添加しなかった場合には、これらのメチル化体の生成は認められなかった。
以上の結果から、エノキタケ培養菌糸体から抽出した粗酵素がメチルトランスフェラーゼ酵素活性を有することが確認された。
【0064】
<各種エノキタケ酵素活性の確認>
エノキタケの採取地の違い等によるメチルトランスフェラーゼ酵素活性の有無を確認した。エノキタケの菌糸は、農業生物資源ジーンバンクより購入したものを使用し、Difco Potato Dextrose Ager培地上で25℃にて培養した。菌糸体の液体培養及び酵素活性測定は、上記<メチルトランスフェラーゼ酵素活性のスクリーニング>と同様にして行った。試験には、MAFF番号430204、430205、430206、430207、430209、435210、430212、430214、430224、435085、430211、430213、435121、440110、440111、及び440118を用いた。その結果、それぞれの菌糸によって酵素活性の強弱はあるものの、全ての菌糸において、メチルトランスフェラーゼ酵素活性を確認した。
【0065】
<エノキタケ由来メチルトランスフェラーゼの同定>
上記と同様にしてエノキタケ菌糸体を液体培養し、得られた菌糸体培養液をろ過して菌糸体を回収し、凍結乾燥した。凍結乾燥物約12gを乳鉢で破砕した後、粗酵素溶解液600mlに懸濁した。この懸濁物を超音波破砕した後、遠心分離(10,000rpm×10min、4℃)し、回収した上清を再度遠心分離(30,000rpm×30min、4℃)して上清を回収した。この上清に、60%飽和になるように硫酸アンモニウムを添加し、攪拌、遠心分離して上清を回収した。さらに、この回収した上清に、80%飽和になるように硫酸アンモニウムを添加し、攪拌、遠心分離して沈殿物を得た。得られた沈殿物を、粗酵素溶解液40mlに溶解した後、PD−10(GEヘルスケアバイオサイエンス社製)を用いて脱塩した。脱塩したサンプルは、トリス緩衝液(20mM Tris−HCl(pH7.5)、1mM DTT)で平衡化した陰イオン交換カラム(HiPrep 16/10 DEAE FF、GEヘルスケアバイオサイエンス社製)に吸着させ、上記トリス緩衝液で調整した0〜500mM NaCl溶液の直線濃度勾配を用いて溶出し、メチルトランスフェラーゼ酵素活性が確認されたフラクションを、活性画分として分取した。なお、メチルトランスフェラーゼ酵素活性の確認は、<メチルトランスフェラーゼ酵素活性のスクリーニング>と同様にして行った。
【0066】
得られた活性画分を、限外ろ過カラムで脱塩、濃縮した後、上記トリス緩衝液で平衡化した陰イオン交換カラム(HiLoad 26/10 Q−sepharose HP、GEヘルスケアバイオサイエンス社製)に吸着させ、上記トリス緩衝液で調整した0〜500mM NaCl溶液の直線濃度勾配を用いて溶出し、活性画分を分取した。得られた活性画分を、限外ろ過カラムにて脱塩、濃縮し、再度同じ陰イオン交換カラムに吸着させ、分取画分を前条件より詳細にして活性画分を分取し、同様に限外ろ過カラムにて脱塩、濃縮を行った。
【0067】
得られた濃縮画分を、150mM NaClを含むトリス緩衝液(20mM Tris−HCl(pH7.5)、1mM DTT、150mM NaCl)で平衡化したゲルろ過カラム(HiLoad 16/60 Superdex 200 prep grade、GEヘルスケアバイオサイエンス社製)を用いて分画した。分取した各画分をSDS−PAGE電気泳動にかけ、得られたタンパク質染色像と、各画分のメチルトランスフェラーゼ酵素活性とを比較したところ、酵素活性と相関したバンドが、約24〜25kDaの位置に確認された。酵素活性が確認された画分を、さらに、上記トリス緩衝液で平衡化した陰イオン交換カラム(TSK−GEL BIOASSIST Q、東ソー社製)に吸着させ、上記トリス緩衝液で調整した0〜500mM NaCl溶液の直線濃度勾配を用いて溶出し、活性画分を分取した。活性画分を、再度SDS−PAGE電気泳動にかけた結果、約24〜25kDaの位置に再度特異的なバンドを確認した。この酵素タンパク質の内部配列を明らかにするために上記陰イオン交換カラム(HiLoad 26/10 Q−sepharose HP、GEヘルスケアバイオサイエンス社製)で精製したサンプルをSDS−PAGE電気泳動にかけ、バンドをゲルから回収した。
【0068】
回収したバンドは、常法に従ってゲル内トリプシン消化処理を行い、LC/MS/MS(サーモフィッシャーサイエンティフィック社製)を用いてタンパク質の内部配列を解析した。さらに、同じ活性画分を二次元電気泳動で電気泳動し(一次元目:pH3〜10の等電点分離、二次元目:4〜20%グラジエントのポリアクリルアミドゲル)、分子量約24〜25kDa、pH5付近のスポットをゲルから回収した。図3は、二次元電気泳動像である。図中、丸で囲んだスポットを、目的の酵素タンパク質として回収した。
【0069】
この回収したスポットに対して常法に従ってゲル内トリプシン消化処理を行い、LC/MS/MS(サーモフィッシャーサイエンティフィック社製)を用いて、スポットに存在しているタンパク質の内部配列の確認を行った。その結果、内部配列RVLEVGTLGGYSTTWLARA(配列番号3)及びTGGIIIVDNVVR(配列番号4)を得た。このアミノ酸配列をもとに、NCBIのデータベースによるホモロジー検索を行った結果、既知のO−メチルトランスフェラーゼの内部配列と高い相同性を有していた。
【0070】
<エノキタケ由来メチルトランスフェラーゼ遺伝子の単離・同定>
上記内部配列の配列情報に基づいてディジェネレートプライマーを設計し、エノキタケから回収したtotal−RNAから、エノキタケ由来メチルトランスフェラーゼ遺伝子を単離・同定した。
具体的には、まず、エノキタケ培養菌糸体約1gを、乳鉢を用いて液体窒素下で破砕したものから、TRI Reagent(シグマ社製)を用いてtotal−RNAを回収した。得られたtotal−RNA約4μgから、サーモスクリプトRT−PCRシステム(インビトロジェン社製)を用いて、55℃、50分間反応させることによりcDNAを合成した。このcDNAを鋳型とし、上記内部配列の配列情報に基づいて設計されたディジェネレートプライマー(FVOMT−F及びFVOMT−R)を用いてPCRを行い、エノキタケ由来メチルトランスフェラーゼ遺伝子の単離を行った。設計されたディジェネレートプライマーの塩基配列及びPCR条件を下記に記す。なお、塩基配列中、Sはグアニン又はシトシンを、Mはアデニン又はシトシンを、Yはチミン又はシトシンを、Dはアデニン、グアニン、又はチミンを、Kはグアニン又はチミンを、Vはアデニン、グアニン、又はシトシンを、Rはグアニン又はアデニンを、それぞれ示す。
【0071】
FVOMT−F:GAGGTSGGMACYYTDGGMGGSTA
FVOMT−R:GCKSACVACRTTRTCMAC
PCR条件:94℃,5min→(94℃,1min→55℃,1min→72℃,1min)×40cycles→72℃,7min
【0072】
PCR産物をアガロースゲル電気泳動にかけた結果、約400bpの増幅バンドが確認された。アガロースゲルから当該バンドを切り出し、QIAquick Gel Extraction Kit(キアゲン社製)を用いてPCR産物を回収し、pGEM−Tベクター(プロメガ社製)へクローニングした後、大腸菌JM−109株(タカラ社製)へ形質転換した。得られた形質転換体を、LB培地にて37℃で終夜振とう培養し、得られた培養物からQIAprep Spin Miniprep Kit(キアゲン社製)を用いてプラスミドを抽出した。抽出されたプラスミドのインサートの塩基配列を、Big Dye Terminator v3.1 Cycle Sequencing Kit(アプライドバイオシステムズ社製)及びABI PRISM 3100−AVANT Genetic Analyzer(アプライドバイオシステムズ社製)により確認した。
【0073】
確認された遺伝子の部分塩基配列をもとに、特異的プライマー(FVOMT−5’GSP1、FVOMT−5’GSP2、FVOMT−3’GSP1、FVOMT−3’GSP2)を設計し、5’側と3’側の全長を取得するためにRACE−PCR法を行った。設計した特異的プライマーの塩基配列を下記に示す。
【0074】
FVOMT−5’GSP1:AGCCTCTTAGCCTCAACAAAGTA
FVOMT−5’GSP2:TCTTCGAGCTCGAAGGTGAT
FVOMT−3’GSP1:ATCACCTTCGAGCTCGAAGA
FVOMT−3’GSP2:TACTTTGTTGAGGCTAAGAGGCT
【0075】
5’RACE−PCRは、まず、単離したtotal−RNA2.5μgから、FVOMT−5’GSP1プライマーを用い、サーモスクリプトRT−PCRシステム(インビトロジェン社製)を用いて、55℃、50分間反応させてcDNAを合成した。次いで、得られたcDNAから、5’RACE System for Rapid Amplification of cDNA Ends,Version 2.0(インビトロジェン社製)を用い、FVOMT−5’GSP1プライマー及びFVOMT−5’GSP2プライマーを用いて5’側の単離を行い、塩基配列を確認した。
同様に、3’RACE−PCRは、サーモスクリプトRT−PCRシステム(インビトロジェン社製)を用いてcDNAを合成し、3’RACE System for Rapid Amplification of cDNA Ends,Version 2.0(インビトロジェン社製)を用いて、得られたcDNAからFVOMT−3’GSP1プライマー及びFVOMT−3’GSP2プライマーを用いて3’側の単離を行い、塩基配列を確認した。
【0076】
確認された5’末端及び3’末端の塩基配列に基づき、下記に示すFVOMT−5’NdeIプライマー及びFVOMT−3’BamHIプライマーを設計し、RT−PCRを行った。得られたPCR産物をアガロースゲル電気泳動し、増幅バンドを回収した。回収したPCR産物をpGEM−Tベクターへクローニングし、大腸菌JM109株へ形質転換後、インサートの塩基配列を確認した。その結果、エノキタケ由来メチルトランスフェラーゼ酵素の全遺伝子を単離するにいたった。同定されたエノキタケ由来メチルトランスフェラーゼ遺伝子の塩基配列は、配列番号1で表される塩基配列であり、配列番号2で表されるアミノ酸配列からなるポリペプチドをコードしている。
【0077】
FVOMT−5’NdeI:TACATATGTCCAACCCGACAAGCATACT
FVOMT−3’BamHI:TAGGATCCAAGTTTGATAGCGTACAAGAATCC
【0078】
[参考例2]
<組換えエノキタケ由来メチルトランスフェラーゼ酵素の製造>
参考例1で単離されたエノキタケ由来メチルトランスフェラーゼ遺伝子を組み込んだ組換え発現ベクターを作製し、この組換え発現ベクターを大腸菌に導入して、大腸菌内で発現させたエノキタケ由来メチルトランスフェラーゼ酵素を回収した。
具体的には、まず、参考例1で作製したエノキタケ由来メチルトランスフェラーゼ遺伝子含有pGEM−Tベクターから、制限酵素NdeIとBamHIで切断し、アガロース電気泳動により、インサートを回収した。このインサートを、pET28a(+)ベクター(ノバジェン社製)のNdeI、BamHIサイトへクローニングすることにより、組換え発現ベクターを作製した。この組換え発現ベクターを大腸菌BL21(DE3)株(ストラタジーン社製)へ導入することにより、当該組換え発現ベクターを含む形質転換体を得た。
【0079】
得られた形質転換体をLB培地にて37℃で終夜振とう培養した後、その一部を再度新しいLB培地へ添加して培養した。O.D.600=0.6付近となるように培養した後、IPTGを最終濃度1mMになるように添加し、さらに28℃で振とう培養して、ヒスチジンタグ付酵素タンパク質の発現誘導を行った。発現誘導した大腸菌を遠心分離し、沈殿物を上記トリス緩衝液に懸濁した。この大腸菌懸濁液を超音波破砕し、再度遠心分離した。得られた上清(ライセート)を、12%SDS−PAGEにて電気泳動した結果、IPTG誘導前の大腸菌を同様に処理して電気泳動した場合と比較して、約29kDa付近に、IPTG処理により発現が誘導されたタンパク質のバンドを確認した。
【0080】
図4は、IPTG処理前の大腸菌のライセート(レーン1)、IPTG処理後の大腸菌のライセート(レーン2)、IPTG処理後の大腸菌のライセートからヒスチジンタグを用いて得られた精製物(レーン3)のSDS−PAGE電気泳動像である。図中、「M」は分子量マーカーを流したレーンである。図中の矢印の位置のバンド(約29kDa)を画像解析し、タンパク質量を数値化して比較した。画像解析には、Photoshop(アドビシステムズ社製)及びScion Image(Scion Corporation社製)ソフトウェアを使用した。その結果、この約29kDaのタンパク質量は、IPTG処理前を0%とした場合、IPTG処理後で8.4%、処理後の精製物で25.1%であり、当該タンパク質が、IPTG処理により発現が誘導されたヒスチジンタグを有するタンパク質であることが確認された。
【0081】
pET28a(+)ベクターのNdeI、BamHIサイトへクローニングした場合、約5kDaのベクター由来発現タンパク質(ヒスチジンタグを含む)が付加される。また、配列番号2のアミノ酸配列から算出された本酵素タンパク質の理論分子量は、24.7kDaである。つまり、図4の矢印で示した約29kDaのタンパク質は、組換え発現ベクターにより導入したアミノ酸配列から得られる推定分子量と一致し、ヒスチジンタグ付きエノキタケ由来メチルトランスフェラーゼ酵素であることが分かった。
【0082】
[参考例3]
<組換えエノキタケ由来メチルトランスフェラーゼ酵素の酵素活性>
参考例2で得られたIPTG処理後の大腸菌のライセートを、エノキタケ由来メチルトランスフェラーゼ酵素の組換え酵素の粗酵素液とし、この粗酵素液のメチルトランスフェラーゼ酵素活性を調べた。
具体的には、20mM Tris−HCl(pH7.5)、2.5mM MgCl2、0.25mM EGCG、0.5mM SAM、及び50% 粗酵素液となるように、酵素反応液3mlを調製し、この酵素反応液を37℃、16時間インキュベートし、反応させた。反応後の酵素反応液を、参考例1の<メチルトランスフェラーゼ酵素活性のスクリーニング>と同様に処理して分析した結果、エピガロカテキン−3−O−(3−O−メチル)ガレート、エピガロカテキン−3−O−(4−O−メチル)ガレート、エピガロカテキン−3−O−(3,4−O−ジメチル)ガレート、エピガロカテキン−3−O−(3,5−O−ジメチル)ガレート、エピ(4−O−メチル)ガロカテキン−3−O−(3,5−O−ジメチル)ガレートの生成が確認できた。
これらの結果から、参考例2において大腸菌内で発現させた組換え酵素が、メチルトランスフェラーゼ酵素活性を有していることが確認された。
【0083】
[参考例4]
<菌糸体抽出粗酵素液を用いたメチル化体の製造>
参考例1と同様にして、エノキタケ菌糸体培養液の菌糸体から抽出し、得られた抽出物の60%〜80%硫酸アンモニウム画分を脱塩処理したものを、エノキタケ由来メチルトランスフェラーゼ酵素の天然型酵素の粗酵素液とした。
具体的には、まず、エノキタケ菌糸体培養液をろ過して菌糸体を回収し、凍結乾燥した。得られた凍結乾燥物を乳鉢にて破砕した後、上記粗酵素溶解液に懸濁して、超音波破砕し、遠心分離して上清を回収した。この上清に、60%飽和になるように硫酸アンモニウムを添加し、攪拌、遠心分離して上清を回収した。さらに、この回収した上清に、80%飽和になるように硫酸アンモニウムを添加し、攪拌、遠心分離して沈殿物を得た。得られた沈殿物を、粗酵素溶解液に溶解した後、PD−10(GEヘルスケアバイオサイエンス社製)を用いて脱塩した。この脱塩サンプルを、エノキタケ由来メチルトランスフェラーゼ酵素の天然型酵素の粗酵素液とした。
得られた粗酵素液の各種基質に対するメチルトランスフェラーゼ活性を確認した。酵素反応液の組成は、20mM Tris−HCl(pH7.5)、2.5mM MgCl2、0.05mM 基質、0.5mM SAM、0.04%(w/v) アスコルビン酸、及び粗酵素液(全量3mlに対して0.25ml)とし、全量3mlを37℃、6時間反応させた。基質には、エピカテキン−3−O−ガレート(ECG)、カテキン−3−O−ガレート(CG)、ガロカテキン−3−O−ガレート(GCG)、カフェ酸、クロロゲン酸、エラグ酸、ブテイン、スルフレチン、ルテオリン、ミリセチン、又はロスマリン酸を、それぞれ用いた。
反応後の酵素反応液を、参考例1<メチルトランスフェラーゼ酵素活性のスクリーニング>と同様に処理して、当該酵素反応液中のメチル化体を、下記の条件でLC−MSを用いて分析した。標品と比較分析が可能なものについては同定も行った。HPLC条件及びMS条件を以下に示す。
【0084】
・HPLC条件
カラム:Inertsil ODS−3、2.1×150mm(GLサイエンス社製)
移動相A:0.1(v/v)%ギ酸水溶液
移動相B:0.1(v/v)%ギ酸を含むアセトニトリル
グラジエント条件(i):8%B液(20分間)→25%B液(88分間)
グラジエント条件(ii):8%B液(10分間)→50%B液(31分間)
流速:0.2ml/min
検出:UV280nm
【0085】
・MS条件
検出器:API3000(アプライドバイオシステムズ社製)
イオン源:ESI(ネガティブ)
カーテンガス:10
ネブライザーガス:14
ターボガス:6l/min
イオンスプレー電圧:−4000V
イオンスプレー温度:500度
コーン電圧:−41V
【0086】
LC−MS分析の結果を表1に示す。この結果、用いた全ての基質に対して、メチル化体の生成が確認された。これらの結果から、本発明の酵素は、水酸基を有する各種化合物を基質とし、メチル基を修飾することが可能であることが明らかとなった。
また、ECGのように、化合物中に複数の水酸基を有する化合物の中には、複数種類のメチル化体の生成が確認され、かつ、ジメチル化体も確認された。この結果から、本発明の酵素は、基質の化合物中の複数の水酸基のそれぞれを独立にメチル化可能であることも確認された。
【0087】
【表1】
【0088】
[参考例5]
<反応時間の検討>
参考例4で用いたエノキタケ由来メチルトランスフェラーゼ酵素の天然型酵素の粗酵素液(以下、天然型酵素)と、参考例2で用いたエノキタケ由来メチルトランスフェラーゼ酵素のヒスチジンタグ精製した組換え酵素液(以下、組換え酵素)とを用いて、酵素反応の反応時間を検討した。
酵素反応液の組成は、20mM リン酸緩衝液(pH7.0)、2.5mM MgCl2、0.05mM EGCG、0.5mM SAM、0.04%(w/v) アスコルビン酸、及び各粗酵素液(全量30mlに対して2.5ml)とし、全量30mlを37℃で反応させた。反応開始から5分、10分、15分、20分、25分、30分、35分、40分、45分、50分、55分、60分、2時間、 4時間、6時間、8時間、24時間後に、酵素反応液を1mlずつ分取した。これらの酵素反応液を、参考例1<メチルトランスフェラーゼ酵素活性のスクリーニング>と同様に処理して、当該酵素反応液中のメチル化体を、下記のHPLC条件で分析した。
【0089】
カラム:Inertsil ODS−3、2.1×150mm(GLサイエンス社製)
移動相A:0.1(v/v)%ギ酸水溶液
移動相B:0.1(v/v)%ギ酸を含むアセトニトリル
グラジエント条件:10%B液(10分間)→16%B液(50分間)
流速:0.2ml/min
検出:UV280nm
【0090】
各反応時間後に分取した酵素反応液中のEGCGメチル化体濃度を図5に示す。図5(A)が天然型酵素の結果であり、(B)が組換え酵素の結果である。この結果、天然型酵素では反応開始8時間後で、組換え酵素では20分後で、EGCGメチル化体の生成が最も高い値を示した。
【0091】
[参考例6]
<最適pHの検討>
参考例5において用いた天然型酵素及び組換え酵素の酵素反応における最適pHを調べた。
酵素反応液の組成は、20mMの各種緩衝液に、2.5mM MgCl2、0.05mM EGCG、0.5mM SAM、0.04%(w/v) アスコルビン酸、及び各粗酵素液(全量3mlに対して0.25ml)となるように添加して調製した。用いた緩衝液は、酢酸緩衝液(pH3.0〜5.5)、リン酸緩衝液(pH6.0〜7.0)、及びTris−HCl(pH7.5〜10)である。調製した酵素反応溶液の全量3mlを、37℃で反応させた。反応時間は、天然型酵素で6時間、組換え酵素で10分間とした。これらの酵素反応液を、参考例1<メチルトランスフェラーゼ酵素活性のスクリーニング>と同様に処理して、当該酵素反応液中のメチル化体を分析した。
各pHの酵素反応液中のEGCGメチル化体濃度を図6に示す。この結果、天然型酵素ではpH6.5〜8.5で、組換え酵素ではpH6以上で、EGCGメチル化体の生成が確認された。
【0092】
[参考例7]
<最適温度の検討>
参考例5で用いた天然型酵素及び組換え酵素の酵素反応における最適温度を調べた。
酵素反応液の組成は、20mM リン酸緩衝液(pH7.0)、2.5mM MgCl2、0.05mM EGCG、0.5mM SAM、0.04%(w/v) アスコルビン酸、及び各粗酵素液(全量3mlに対して0.25ml)とし、全量3mlを、4、10、20、30、37、42、50、又は60℃で、それぞれ反応させた。反応時間は、天然型酵素で6時間、組換え酵素で10分間とした。これらの酵素反応液を、参考例1<メチルトランスフェラーゼ酵素活性のスクリーニング>と同様に処理して、当該酵素反応液中のメチル化体を分析した。
各温度における酵素反応液中のEGCGメチル化体濃度を図7に示す。この結果、天然型酵素では10〜50℃において、組換え酵素では行った全ての温度において、EGCGメチル化体の合成が確認された。中でも、30〜42℃で、両酵素のいずれにおいても良好な酵素活性が確認された。
【0093】
[参考例8]
<基質となるテアフラビン標準品の調製>
島根県出雲産の紅茶(商品名:出雲国産紅茶(リーフ)、西製茶所製)を粉砕し、40倍量の2%(w/v)アスコルビン酸含有50%(v/v)EtOH水溶液を用いて、20分間の室温抽出を3回行った。抽出液は遠心分離を行い、抽出液から茶葉を取り除いた。抽出液は、減圧濃縮により有機溶媒を除去した後、Sep−Pak Vac C18(ウォーターズ社製)に供し、15%、40%、100%EtOHステップワイズ溶出を行った。テアフラビンが含まれる40%EtOH画分を凍結乾燥させ、適宜40%EtOHに再溶解し、下記の条件によりテアフラビンの分取を行った。分取したテアフラビンはSep−Pak Vac C18を用いて脱酸し、凍結乾燥品を作製した。これにより、TF3G標準品、TF3’G標準品、TF3,3’G標準品が得られた。
【0094】
分取条件;
カラム:Inertsil ODS−3(20 x 250mm、GLサイエンス社製)移動相A:0.05(v/v)%リン酸水溶液
移動相B:12.5(v/v)%酢酸エチルを含むアセトニトリル
グラジエント条件:24%B液(50分間)→50%B液(0.1分間)→50%B液(10分間)
流速:18ml/min
カラム温度:30℃
検出:UV280nm
【0095】
[実施例1]
<メチル化テアフラビンの製造>
参考例2において製造した組換え酵素を用いて、メチル化テアフラビンの製造を行った。基質としては、参考例8で得られたTF3G標準品、TF3’G標準品、TF3,3’G標準品をそれぞれ用いた。
酵素反応液の組成は、20mMリン酸緩衝液(pH6.5)、2.5mM MgCl2、0.025%(w/v)テアフラビン(基質)、0.5mM SAM、0.04%(w/v)アスコルビン酸、及びメチルトランスフェラーゼ(全量1000mlに対して33.3ml)とし、37℃、6時間で反応させた。その後、酵素反応液1000mlに対し、24mlの1N HClを加え、酵素反応を終了させた。酵素反応液に等量の酢酸エチルを加えて激しく撹拌し、静置後、酢酸エチル層を回収した。残る水層に対し、再度同様の操作を行った。回収した酢酸エチル層は減圧濃縮した後、参考例8と同じ条件でメチル化テアフラビンの分取を行った。分取したメチル化テアフラビンは、Sep−Pak C18を用いて脱酸し、凍結乾燥品を作製した。得られたメチル化テアフラビン凍結乾燥品に対して下記の条件でTOF−MS分析を行い、得られた精密分子量から分子式を算出した。また、下記の条件でNMR(H−NMR、C−NMR、HSQC、HMBC、COSY)を測定し、構造同定を行った。
【0096】
TOF−MS条件;
検出器:QSTAR ELITE(アプライドバイオシステムズ社製)
イオン源:ESI(ネガティブモード)
カーテンガス:30
イオンソースガス1:50
イオンソースガス2:50
イオンスプレー電圧:−4500V
イオンスプレー温度:450℃
デクラスタリングポテンシャル:−30V
フォーカスポテンシャル:−250
デクラスタリングポテンシャル:−15V
【0097】
NMR条件;
検出器:XWIN−NMR AV600(ブルカーバイオスピン社)
プローブ温度:300K
溶媒:アセトン−d6
【0098】
TOF−MS、及びNMR解析の結果を表2〜4に示した。既知化合物である3MeTF3Gの他に8種の新規化合物が得られた。各新規化合物をそれぞれ、3,5diMeTF3G(化学式(I)−1)、3MeTF3’G(化学式(II)−1)、4MeTF3’G(化学式(II)−2)、3,5diMeTF3’G(化学(II)−3)、3,4diMeTF3’G(式(II)−4)、3MeTF3,3’G(化学式(III)−1)、4MeTF3,3’G(化学式(III)−2)、3,5diMeTF3,3’G(化学式(III)−3)と決定した。
【0099】
【表2A】
【0100】
【表2B】
【0101】
【表3A】
【0102】
【表3B】
【0103】
【表3C】
【0104】
【表4A】
【0105】
【表4B】
【0106】
[実施例2]
<メチル化テアフラビン含有紅茶ポリフェノールの製造、及び反応効率>
標準品に代えて参考例8において調製した40%EtOH画分の凍結乾燥品を基質として、実施例1と同様に酵素反応を行った。酵素反応液は経時的に0.5mlずつ取り分け、実施例1と同様に処理した後、下記の条件で分析することにより、テアフラビンとメチル化テアフラビンの組成を調べた。
【0107】
分析条件;
カラム:Inertsil ODS−3(4.6 x 250mm、GLサイエンス社製)移動相A:0.05(v/v)%リン酸水溶液
移動相B:12.5(v/v)%酢酸エチルを含むアセトニトリル
グラジエント条件:24%B液(50分間)→50%B液(0.1分間)→50%B液(10分間)
流速:1ml/min
カラム温度:50℃
検出:UV375nm
【0108】
メチル化テアフラビン含有紅茶ポリフェノールの製造過程において、テアフラビンとメチル化テアフラビンの組成の推移を経時的に調べた結果を図8に示す。この結果、反応時間の経過とともにメチル化テアフラビンの生成量は増加し、24時間後で生成量が最大になることが明らかになった。この結果から、メチルトランスフェラーゼを用いた酵素処理によって、紅茶ポリフェノール中のメチル化テアフラビン含量を増やすことが可能であることが明らかとなった。
【0109】
[実施例3]
<メチル化テアフラビンのpH安定性評価>
実施例1で用いたテアフラビン標準品及び実施例1で得られたメチル化テアフラビンを、400μMになるように、40%(v/v)エタノール含有0.3M 酢酸緩衝液(pH5.0)又は40%(v/v)エタノール含有0.3M Tris−HCl緩衝液(pH7.5)にそれぞれ溶解し、常温におけるpH安定性を調べた。反応開始0、1、2時間後に試料を50μlずつ採取し、等量の1%(w/v)アスコルビン酸水溶液を添加して分解反応を停止させ、 40%(v/v)になるようにEtOHを加え、残存する各種テアフラビンを実施例2に記載の方法で分析した。
【0110】
反応開始2時間後のメチル化テアフラビン残存率を図9に示す。図9(A)がpH5.0の結果であり、図9(B)がpH7.5の結果である。この結果、ポリフェノールが安定とされるpH5.0では、全ての化合物がほぼ100%残存していた。一方、ポリフェノールが不安定とされるpH7.5では、メチル化テアフラビンはテアフラビンよりも良好なpH安定性を有していた。また、評価した全ての化合物において、メチル基の数が増えるほど、pH安定性が高まることが明らかになった。
【0111】
[実施例4]
<メチル化テアフラビンのヒト肝組織由来S9フラクションを用いた代謝安定性試験>
ヒト肝組織由来のS9フラクション(シグマ アルドリッチ社製)を用いて、実施例1で用いたテアフラビン標準品及び実施例1で得られたメチル化テアフラビンの代謝安定性を評価した。
反応組成は50mM Tris−HCl緩衝液(pH7.5)、0.1mM 3’−ホスホアデノシン−5’−ホスホサルフェート、1mM ウリジン−5’二リン酸−α−D−グルクロン酸、1mM ニコチンアミドアデニンジヌクレオチドリン酸、10mM MgCl2、0.063% ウシ血清アルブミン、8mM DTT、ヒト肝組織由来S9フラクション(最終量50μgタンパク質)、5μM 各種テアフラビン又はメチル化テアフラビンからなる反応液250μlを調製し、37℃でインキュベートした。反応開始0、15、30、60分後に50μlずつ分取し、45μlの氷冷したメタノールと、5μlの1%(w/v)アスコルビン酸水溶液とを加えて攪拌した。その後、14,000gで3分間遠心分離し、得られた上清から90μlを回収し、残存する各種テアフラビンを実施例2に記載の方法で分析した。
【0112】
反応開始60分後の、各種テアフラビン残存率を図10に示す。その結果、いずれのメチル化テアフラビンも、対応するテアフラビンと比較して残存率が高く、良好な代謝安定性を示した。また、評価した全ての化合物において、メチル基が増えるほどその代謝安定性が向上することが明らかになった。
【0113】
[実施例5]
<メチル化テアフラビンの脂肪蓄積抑制作用評価>
TF3Gのメチル化体の脂肪蓄積抑制作用を調べた。Tea extract(参考例8において、Sep−Pak Vac C18に供する前の抽出液)、実施例1で用いたTF3G(標準品)、実施例1で得られた3MeTF3G及び3,5diMeTF3Gを被検素材とした。
【0114】
まず、ヒト臓器脂肪由来前駆脂肪細胞(Lot.No.8F3482,三光純薬社製)を、前駆脂肪細胞用増殖培地で37℃、5%CO2インキュベーターにて培養し、増殖させた。その後、96−well plate(住友ベークライト社製)に前駆脂肪細胞を1.5×104 cells/wellとなるように撒いて24時間培養した後、培地を前駆脂肪細胞用分化培地に交換して分化を誘導した。分化誘導と同時に、最終濃度がTea extractは10μg/ml、各被検素材は1又は3μMとなるようにDMSO(ジメチルスルホキシド)に溶解して添加した。その後、1日おきに、前駆脂肪細胞用分化培地にTea extractは10μg/ml、各被検素材は1又は3μMとなるように含有させた培地に交換し、10日間培養した。培養終了後、細胞内の脂肪蓄積量をAdipoRedassay reagent(Lonza Walkersville)を用いて蛍光強度により評価した(Ex:485nm, Em:572nm)。
【0115】
各処理を施した細胞群の脂肪蓄積量の測定結果を図11に示す。図11(A)は各被検素材を1μMとなるように培地中に添加した場合の結果を、図11(B)は3μMとなるように培地中に添加した場合の結果を、それぞれ示す。この結果、TF3G、3MeTF3G、又は3,5diMeTF3Gを添加した場合には、無添加の場合やTea extractを添加した場合よりも数値が低く、脂肪蓄積が抑制されていることが明らかとなった。また、3MeTF3G及び3,5diMeTF3Gは、非メチル化体であるTF3Gよりも脂肪蓄積抑制作用が強く、中でも、3,5diMeTF3Gは、既知化合物である3MeTF3Gよりも優れた脂肪蓄積抑制作用を有することが明らかとなった。
これらの結果から、テアフラビンのうち、少なくともTF3Gにおいては、メチル化されている水酸基の数が多くなるほど、脂肪蓄積抑制作用が高まることが明らかになった。
【0116】
[実施例6]
<メチル化テアフラビンの抗酸化作用評価>
ヒト肝癌細胞株HepG2への過酸化水素水による酸化ストレスに対する、メチル化テアフラビンの抑制効果を評価した。Tea extract(参考例8において、Sep−Pak Vac C18に供する前の抽出液)、実施例1で用いたTF3’G及びTF3,3’G(標準品)、並びに実施例1で得られた3MeTF3’G、3,5diMeTF3’G、3MeTF3,3’G、4MeTF3,3’G、及び3,5diMeTF3,3’Gを被検素材とした。これらの被検素材は、予めDMSOに溶解させた。
【0117】
HepG2細胞は、10%非働化FBS(ウシ胎児血清)、100unit/ml ペニシリン、100μg/mlストレプトマイシン含有D−MEM培地で、37℃、5%CO2インキュベーターにて継代培養した。その後、96−well plate(住友ベークライト社製)にHepG2細胞を4×104 cells/wellとなるように撒き、最終濃度がTea extractは0.6、6、60μg/ml、被検素材は0.2、2、又は20μMとなるようにDMSOに溶解して添加したD−MEM培地で、37℃、5%CO2インキュベーターで23時間培養した。培地を除去した後、各ウェルに150μlの12mM 過酸化水素水含有D−MEM培地を添加し、細胞を2時間、過酸化水素水に暴露した。その後、上清を除去した各ウェルに、200μlの10%アラマーブルー含有D−MEM培地を添加し、3時間培養後、蛍光強度を測定した(Ex:544nm, Em:590nm)。
【0118】
過酸化水素水を添加しなかった場合のHepG2細胞生存率を100%とし、各濃度の被検素材を添加した場合におけるHepG2細胞生存率を求めた。HepG2細胞生存率の測定結果を図12に示す。図12(A)がTF3’G及びそのメチル化体を添加した場合の測定結果であり、図12(B)がTF3,3’G及びそのメチル化体を添加した場合の測定結果である。なお、有意差の検定はTukey法を用いた。
【0119】
過酸化水素水のみを添加した場合やTea extractを添加した場合と比較した結果、テアフラビンを添加した場合に差は確認できなかったものの、メチル化テアフラビンを添加した場合は細胞生存率がほぼ100%であり、有意に酸化ストレスが抑制されることが判明した。特に、TF3’G及びTF3,3’Gにおいては、メチル化テアフラビンのほうが、非メチル化体よりも高い抗酸化作用を有することが確認された。
なお、各被検素材を2又は20μMとなるように添加した場合には、非メチル化体と各種メチル化体の間で有意な差は得られなかった。これは、過酸化水素水濃度等の処理条件により、抗酸化作用の効果が必要十分であったためと推察される。
【0120】
なお、参考例1〜8及び実施例1〜6において使用した試薬の購入先は、以下の通りである。
Difco Potato Dextrose Ager:日本ベクトン・ディッキンソン
Peptone: 日本ベクトン・ディッキンソン
Yeast Extract:日本ベクトン・ディッキンソン
前駆脂肪細胞用増殖培地:三光純薬
D−MEM培地:和光純薬
分化培地:三光純薬
アラマーブルー:インビトロジェン
FCS:JRHバイオサイエンス
ペニシリン:GIBCO
ストレプトマイシン:GIBCO
EGCG:テアビゴ、DSMニュートリションジャパン
【0121】
エピカテキン−3−O−ガレート(ECG):フナコシ
カテキン−3−O−ガレート(CG):フナコシ
ガロカテキン−3−O−ガレート(GCG):フナコシ
カフェ酸:シグマ
クロロゲン酸:フナコシ
エラグ酸:フナコシ
ブテイン:フナコシ
スルフレチン:フナコシ
ルテオリン:フナコシ
ミリセチン:フナコシ
ロスマリン酸:フナコシ
【産業上の利用可能性】
【0122】
本発明の新規メチル化テアフラビンは、安定性及び機能性に優れているため、飲食品、医薬品、医薬部外品、化粧品等の様々な商品の製造分野において利用が可能である。
【特許請求の範囲】
【請求項1】
1又は2以上の水酸基がメチル化されたガロイル基を有するメチル化テアフラビンであって、下記一般式(I)〜(III)のいずれか1の式で表されることを特徴とする化合物。
【化1】
[式(I)中、R1〜R3は水素原子又はメチル基を表し、かつ、R1〜R3の少なくとも2以上がメチル基である。]
【化2】
[式(II)中、R4〜R6は水素原子又はメチル基を表し、かつ、R4〜R6の少なくとも1以上がメチル基である。]
【化3】
[式(III)中、R7〜R12は水素原子又はメチル基を表し、かつ、R7〜R12の少なくとも1以上がメチル基である。但し、R7のみがメチル基である場合を除く。]
【請求項2】
下記式(I)−1、(II)−1〜4、及び(III)−1〜3からなる群より選択される1の式で表されることを特徴とする、請求項1に記載の化合物。
【化4】
【化5】
【化6】
【請求項3】
請求項1又は2に記載の化合物を含むことを特徴とする組成物。
【請求項4】
請求項1又は2に記載の化合物を含むことを特徴とする脂肪蓄積抑制剤。
【請求項5】
前記化合物が式(I)−1で表される化合物であることを特徴とする請求項4記載の脂肪蓄積抑制剤。
【請求項6】
請求項1又は2に記載の化合物を含むことを特徴とする抗酸化剤。
【請求項7】
前記化合物が、式(II)−1、(II)−3、(III)−1、(III)−2、及び(III)−3で表される化合物から選択される1種以上であることを特徴とする請求項6記載の抗酸化剤。
【請求項8】
請求項1又は2に記載の化合物を含むことを特徴とする飲食品。
【請求項9】
請求項1又は2に記載の化合物を含むことを特徴とする医薬品又は医薬部外品。
【請求項10】
請求項1又は2に記載の化合物を含むことを特徴とする化粧品。
【請求項11】
下記式(I)−0で表される化合物、下記式(II)−0で表される化合物、及び下記式(III)−0で表される化合物からなる群より選択される1種以上の化合物を基質として、メチルトランスフェラーゼ酵素を反応させることにより、下記一般式(I)で表される化合物、下記一般式(II)で表される化合物、及び下記一般式(III)で表される化合物からなる群より選択される1種以上の化合物を合成することを特徴とする、メチル化テアフラビンの製造方法。
【化7】
【化8】
[式(I)中、R1〜R3は水素原子又はメチル基を表し、かつ、R1〜R3の少なくとも2以上がメチル基である。]
【化9】
[式(II)中、R4〜R6は水素原子又はメチル基を表し、かつ、R4〜R6の少なくとも1以上がメチル基である。]
【化10】
[式(III)中、R7〜R12は水素原子又はメチル基を表し、かつ、R7〜R12の少なくとも1以上がメチル基である。但し、R7のみがメチル基である場合を除く。]
【請求項12】
前記メチルトランスフェラーゼ酵素が、エノキタケ(Flammulina velutipes)由来の酵素であることを特徴とする請求項11記載のメチル化テアフラビンの製造方法。
【請求項1】
1又は2以上の水酸基がメチル化されたガロイル基を有するメチル化テアフラビンであって、下記一般式(I)〜(III)のいずれか1の式で表されることを特徴とする化合物。
【化1】
[式(I)中、R1〜R3は水素原子又はメチル基を表し、かつ、R1〜R3の少なくとも2以上がメチル基である。]
【化2】
[式(II)中、R4〜R6は水素原子又はメチル基を表し、かつ、R4〜R6の少なくとも1以上がメチル基である。]
【化3】
[式(III)中、R7〜R12は水素原子又はメチル基を表し、かつ、R7〜R12の少なくとも1以上がメチル基である。但し、R7のみがメチル基である場合を除く。]
【請求項2】
下記式(I)−1、(II)−1〜4、及び(III)−1〜3からなる群より選択される1の式で表されることを特徴とする、請求項1に記載の化合物。
【化4】
【化5】
【化6】
【請求項3】
請求項1又は2に記載の化合物を含むことを特徴とする組成物。
【請求項4】
請求項1又は2に記載の化合物を含むことを特徴とする脂肪蓄積抑制剤。
【請求項5】
前記化合物が式(I)−1で表される化合物であることを特徴とする請求項4記載の脂肪蓄積抑制剤。
【請求項6】
請求項1又は2に記載の化合物を含むことを特徴とする抗酸化剤。
【請求項7】
前記化合物が、式(II)−1、(II)−3、(III)−1、(III)−2、及び(III)−3で表される化合物から選択される1種以上であることを特徴とする請求項6記載の抗酸化剤。
【請求項8】
請求項1又は2に記載の化合物を含むことを特徴とする飲食品。
【請求項9】
請求項1又は2に記載の化合物を含むことを特徴とする医薬品又は医薬部外品。
【請求項10】
請求項1又は2に記載の化合物を含むことを特徴とする化粧品。
【請求項11】
下記式(I)−0で表される化合物、下記式(II)−0で表される化合物、及び下記式(III)−0で表される化合物からなる群より選択される1種以上の化合物を基質として、メチルトランスフェラーゼ酵素を反応させることにより、下記一般式(I)で表される化合物、下記一般式(II)で表される化合物、及び下記一般式(III)で表される化合物からなる群より選択される1種以上の化合物を合成することを特徴とする、メチル化テアフラビンの製造方法。
【化7】
【化8】
[式(I)中、R1〜R3は水素原子又はメチル基を表し、かつ、R1〜R3の少なくとも2以上がメチル基である。]
【化9】
[式(II)中、R4〜R6は水素原子又はメチル基を表し、かつ、R4〜R6の少なくとも1以上がメチル基である。]
【化10】
[式(III)中、R7〜R12は水素原子又はメチル基を表し、かつ、R7〜R12の少なくとも1以上がメチル基である。但し、R7のみがメチル基である場合を除く。]
【請求項12】
前記メチルトランスフェラーゼ酵素が、エノキタケ(Flammulina velutipes)由来の酵素であることを特徴とする請求項11記載のメチル化テアフラビンの製造方法。
【図1】


【図2】

【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図3】
【図4】
【図2】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図3】
【図4】
【公開番号】特開2011−256165(P2011−256165A)
【公開日】平成23年12月22日(2011.12.22)
【国際特許分類】
【出願番号】特願2011−108449(P2011−108449)
【出願日】平成23年5月13日(2011.5.13)
【出願人】(000000055)アサヒグループホールディングス株式会社 (535)
【Fターム(参考)】
【公開日】平成23年12月22日(2011.12.22)
【国際特許分類】
【出願日】平成23年5月13日(2011.5.13)
【出願人】(000000055)アサヒグループホールディングス株式会社 (535)
【Fターム(参考)】
[ Back to top ]